
Укажите что воспринимают механорецепторы кожи
Обновлено: 29.04.2024
Механорецепторы (греч. mechane орудие, машина + рецепторы) — специальные чувствительные нервные окончания (рецепторы), приспособленные для восприятия механических раздражений. К Механорецепторам относятся рецепторы органов слуха, восприятия гравитации, вестибулярного аппарата, сердечно-сосудистой системы, внутренних органов, кожи, опорно-двигательного аппарата. В зависимости от локализации и функции ряд Механорецепторов получил самостоятельное название — напр., рецепторы опорно-двигательного аппарата — проприоцепторы (см.), рецепторы сосудов — ангиоцепторы (см.), рецепторы, воспринимающие вибрацию,— виброрецепторы (см. Вибрационная чувствительность) и т. д.
В любом Механорецепторе различают: вспомогательный аппарат, механические свойства к-рого, а также и свойства окружающих тканей обеспечивают особенности проведения механического стимула (раздражения) к рецептору; собственно рецепторный субстрат и связанную с ним систему генерации локальных электрических явлений (рецепторного потенциала); структуры, ответственные за генерацию импульсной активности, на основе к-рых ц. н. с. получает сведения о характере действующего стимула (раздражения). Характерной особенностью большинства М. является их чувствительность к направлению действия раздражителя (дирекционная чувствительность), к-рая может быть обусловлена как особенностями строения вспомогательных структур, так и строением самих рецепторных (волосковых) клеток или нервных окончаний (см.). При одном направлении действия раздражителя в рецепторах возникают деполяризационный потенциал и импульсная активность, при другом — гиперполяризационный потенциал, препятствующий возникновению спонтанной импульсной активности. Деятельность практически всех М. находится под контролем ц. н. с., к-рая оказывает свое регулирующее действие как прямым (нервным), так и опосредованным (гуморальным) путем, что изменяет свойства вспомогательных структур, самих рецепторных клеток и отходящих от них афферентных волокон (см. Нервная регуляция функций, Нейрогуморальная регуляция). Большую роль в деятельности многих М. играют свойства окружающей их среды. В жидкости каналов улитки внутреннего уха выявлена высокая концентрация ионов калия (в эндолимфе перепончатого канала улитки человека она, напр., в 30 раз больше, чем в плазме крови) и сниженное содержание ионов натрия. Считается, что повышение концентрации ионов калия в окружающей среде и действие стационарной гиперполяризации М. способствует повышению их чувствительности.

Схематическое изображение строения волосково-реснитчатого (I) и ареснитчатого (II) механорецепторов позвоночных животных; стрелками обозначено направление нейрогуморальной регуляции активности рецепторов: соматическая эфферентная иннервация элементов ткани (кровеносные сосуды, мышечные волокна) — зеленые стрелки, вспомогательного аппарата (мышцы наружного уха, интрафузальные волокна) — красные стрелки; вегетативная эфферентная иннервация элементов ткани, вспомогательного аппарата и окружающих сосудов — оранжевые стрелки; гуморальные вещества, выделяющиеся из сосудов, — прерывистые стрелки ( красные — возбуждающего действия, синие — тормозного действия). Стрелки в синапсе и афферентном нервном волокне показывают направление процессов возбуждения (красные) и торможения (синие). Внешнее воздействие (стимул) — фиолетовые стрелки.
Несмотря на разнообразие Механорецепторов, обусловленное прежде всего особенностями их вспомогательных структур, связанными со спецификой иннервируемых тканей и органов, все они могут быть подразделены на два основных типа. М. первого типа обладают специализированными волосково-реснитчатыми структурами (так наз. волосково-реснитчатые М.), участвующими в актах первичной рецепции (см.). М. второго типа лишены таких образований (ареснитчатые М.). Восприятие раздражителя в них осуществляется непосредственно механочувствительной мембраной нервного окончания. У позвоночных животных волосково-реснитчатый комплекс находится в специальной волосковой клетке, к-рая несет собственно-рецепторную функцию. Волосково-реснитчатые М. обладают высокой чувствительностью к действию адекватного механического раздражителя (напр., М. органов слуха способны воспринимать столь малые смещения, как доли ангстрема). Регулирование их деятельности со стороны ц. н. с. является исключительно тормозным (цветн. рис., I). Ареснитчатые М. позвоночных менее чувствительны, и эфферентные влияния на них преимущественно носят облегчающий характер или даже возбуждающий, как, напр., в случае мышечных веретен (цветн. рис., II). Структурное и функц, разнообразие М. обусловливает большие различия в методах их исследования, использующих как психофизические, так и многочисленные современные аналитические (Электрофизиологические, биофизические и т. д.) подходы.
Патология Механорецепторов весьма разнообразна и тесно связана с патологией тканей и органов, в к-рых они заложены, и наблюдается, напр., при вибрационной болезни (см.), нек-рых видах глухоты (см.), расстройствах вестибулярной функции (см. Вестибулярный симптомокомплекс), атеросклеротических поражениях сердечно-сосудистой системы и т. д. При этом патологические изменения, затрагивающие структуры вспомогательного аппарата М. первого типа (волосково-реснитчатых М.), обратимы и могут поддаваться лечению (напр., при ухудшении деятельности звукопроводящих структур органа слуха). Повреждения же собственнорецептирующих структур носят необратимый характер и не поддаются лечению. Разрушения рецептирующих структур и афферентных нервных волокон М. второго типа (ареснитчатых М.) являются обратимыми и поддаются лечению, напр, возможно восстановление тактильной и проприоцептивной чувствительности вследствие мероприятий, направленных на регенерацию поврежденных афферентных проводников.
Библиография: Гранит Р. Электрофизиологическое исследование рецепции, пер. с англ., М., 1957; он же, Основы регуляции движений, пер. с англ., М., 1973; Ильинский О. Б. Физиология сенсорных систем, ч. 3. Физиология механорецепторов, Л., 1975; Физиология сенсорных систем, под ред. А. С. Батуева, Л., 1976.

Рецепторный аппарат сформирован в результате эволюции и в человеческом организме представлен очень широко.
Механорецепция позволяет нам осязать предметы, различать температурные, вибрационные характеристики, представлять свое расположение в пространстве.
По этой причине их называют тактильными рецепторами кожи.
Что такое рецепторы?
В переводе с латинского языка глагол «recipere» означает «принимать». В этом и заключена биологическая роль рецепторов – прием, обработка и преобразование полученной информации. С точки зрения эволюции — это очень важное приспособление. Ведь среди многочисленных раздражителей очень сложно вычленить и обработать нужные стимулы, обладая тонко организованной нервной системой, способной работать только с потенциалами действия.
Рецепторы являются важной составной частью анализаторных систем. Например, зрение. Первичное и едва ли не самое важно звено этой системы – так называемые палочки и колбочки сетчатки. По морфологии — это видоизмененные нервные клетки. А по своей биологической и физиологической сути — это рецепторы. Именно в них первично происходит восприятие раздражения и первичная обработка полученной информации из внешней среды.
Согласно классификации, рецепторы могут обрабатывать импульсы с поверхностей внутренних органов, стенок их слизистых. Это внутренние рецепторы (механорецепторы легких или, например, пластинчатые тельца поджелудочной железы).
Экстерорецепторы – следующая группа клеток, которые кодируют информацию, полученную извне. К ним относят механорецепторы. Стимулы – давление на рецепторные участки, их деформация либо смещение.
Функции механорецепторов и их виды
Как извне, так и изнутри организм человека испытывает воздействие различного рода раздражителей. Механорецепторы – одна из самых важных групп рецепторов, включающая разнородные клетки. Они преобразуют механические факторы в унифицированный нервный импульс с формированием потенциала действия.
Механорецепторы возбуждаются при действии следующих факторов:
- свет;
- давление;
- сжатие;
- температура;
- звуковые волны;
- вибрация.
Таким образом, суть работы описываемых клеточных структур любого анализатора сводится к следующим важным процессам:
- Раздражение клетки-рецептора различными стимулами.
- Преобразование энергии импульса извне или изнутри в потенциал действия.
- Так называемая эфферентная регуляция.
Среди механорецепторов выделяют первичночувствующие рецепторы. Все описанные выше процессы протекают в пределах одной и той же морфологической структуры (клетки). К ним можно отнести сердечную мускулатуру, эндотелиальные клетки сосудистой выстилки, рецепторы, расположенные в эпидермальном слое кожного покрова. Это очень распространенная группа.
Выделяют также вторичночувствующие рецепторы. К ним относят механорецепторы слухового и вестибулярного анализаторов. Раздражение происходит в одной клеточной структуре, в то время как формирование потенциала действия замыкается на другой.
Далее речь пойдет о клетках первого типа, ведь группа тактильных механорецепторов представлена намного шире.

Тельца Мейснера
Иное название этой гистологической структуры – осязательное тельце. Оно воспринимает механические колебания с частотой диапазона 30000-40000 Герц.
Морфология этого механорецептора несложная. Извне от дермы осязательное тельце отделено капсулой из соединительной ткани. Внутри расположены в форме зигзага безмиелиновые ветви дендритов нервных клеток. На них расположены глиоциты, не покрытые миелином. Относительно оси тельца Мейснера глиальные клетки расположены под прямым углом.

Тельца Мейснера реагируют на вибрацию, давление именно посредством глиоцитов. На последние действуют указанные механические факторы, затем раздражение передается на дендриты нервных клеток. По этим отросткам потенциал действия доходит до следующих нейронов, формируя в конечном итоге ощущение давления (осязание) и вибрации.
Осязательные тельца Мейснера встречаются в следующих органах и тканях человеческого организма:
- сосочковый слой дермы кожного покрова пальцев, подошв;
- кожа области век;
- ареолы сосков, а также сами соски;
- красная кайма губ;
- слизистая оболочка половых органов (большое скопление в так называемой точке G).
Распространенность и плотность телец Мейснера высока. Расположение относительно поверхности кожи перпендикулярное.
Тельца Меркеля
Эти клеточные структуры расположены несколько поверхностнее, чем тельца Мейснера. Локализация – базальный, шиповатый слои эпидермального покрова кожи. На поверхности выстилки волосяных луковиц их также достаточно много. Есть данные, что клетки Меркеля в эпидермисе безволосой части кожи представлены гораздо чаще, больше. На волосистой части головы, к примеру, их не так много.
Клетка содержит разветвляющиеся выросты в виде пальцев, они вплетаются в окружающие ее клеточные и тканевые структуры. Эта особенность позволяет воспринимать сигналы (легкое прикосоновение) с большой площади кожного покрова.
Иначе описываемые рецепторы называются дисками Меркеля (из-за визуальной схожести). Совокупность нескольких телец — это так называемая тактильная корпускула. В ней содержится до 50 дискоидных структур.
Происхождение клеток Меркеля – вопрос спорный. Чаще всего морфологи и цитологи сходятся на том, что это производные нейроэндокринной системы (APUD). Функция их заключена в участии в формировании тактильных ощущений (на легкое статическое прикосновение). Кроме того, учитывая происхождение, они стимулируют питание нервных волокон кожного покрова и его дериватов.
Тельца Руффини
Эта разновидность механорецепторов позволяет воспринимать чувство прикосновения. Расположение – папиллярный слой дермы. Кроме того, их можно обнаружить в поверхностных слоях подкожно-жировой клетчатки и жировой ткани.
По морфологии тельца Руффини напоминают колбу. Сердцевина заполнена безмиелиновыми волокнами, расположенными в основном спиралевидно. Следом идет пространство капсулы, отграниченное от внутренней колбы мембраной. В нем сосредоточены клеточные и волокнистые структуры, характерные для соединительной ткани. Они выполняют трофическую функцию для основных элементов сердцевины тельца Руффини.
Капсульное пространство заполнено межклеточной жидкостью. Следующий компонент — соединительнотканная капсула. Она имеет слоистое строение, почему этот рецептор иногда называют луковицей Руффини. Количество слоев – около 4-5. Они состоят из четко ориентированных фиброцитов.
Кроме растяжения, позволяют принимать температурные импульсы. Это оправдывает их расположение в нижних слоях эпидермиса, в сосочковом слое дермы и подкожно-жировой клетчатке. Посредством работы луковиц Руффини происходит восприятие прикосновения, тепла.

Тельца Фаттера-Пачини
Иное название этой разновидности клеточных структур – пластинчатые тельца. Это связано с морфологическими особенностями. Установлено, что тельца Фатера-Пачини являются рецепторами, реагирующими на вибрационные раздражители.
Внешне эти структуры напоминают луковицу. Это связано со слоистым расположением соединительной ткани, окружающей нервные безмиелиновые окончания. Между слоями находится жидкость, по свойствам напоминающую ликвор. Функция тельца Пачини – реакция на давление и вибрацию.

Формирование нервного импульса является результатом сжатия клетки, которое вызывает скольжение слоев капсулы тельца друг относительно друга. Это раздражение улавливается нервными окончаниями и передается по афферентным путям на нейроны 1 уровня.
Колбы Краузе
В отличие от остальных тактильных механорецепторов они представлены сферическими клетками. По сути, это разветвления дендритов нервных клеток кожного покрова, свернутые в клубочковую структуру. Они окружены соединительной тканью.
Большое количество этих рецепторов сосредоточено на слизистой оболочке полости рта, языка. Замечено, что колбы Краузе находятся также на слизистой оболочке женских половых органов.
Кроме тактильной чувствительности, эти рецепторы ответственны за холодовые ощущения.

Механорецепторы позволяют реагировать на прикосновение, щекотание, вибрацию и осязание. Они обрабатывают поступающие сигналы. В итоге мы имеем представление о своем расположении, о свойствах окружающих предметов и собственного тела.
Соматовисцеральная система перерабатывает сенсорную информацию нескольких модальностей, получая ее от кожи, мышц, суставов, внутренних органов и кровеносных сосудов. Она содержит разные типы рецепторов: механорецепторы, терморецепторы, хеморецепторы, осморецепторы, но-цицепторы. Поступающая информация передается по афферентным волокнам в составе чувствительных и смешанных нервов, перерабатывается раздельными проводящими путями и переключательными ядрами, а затем доставляется в соматосенсорную кору, занимающую постцентральные извилины мозга и организованную топически.
Тактильная чувствительность
Часть соматовисцеральной системы, обеспечивающая чувство осязания, включает несколько разновидностей механорецепторов кожи, представленных свободными нервными окончаниями либо инкапсулированными, т. е. заключенными в капсулу из соединительной ткани или видоизмененных клеток эпидермиса (рис. 17.4). Свободные нервные окончания иннервируют волосяные фолликулы пушковых волос, покрывающих большую часть тела человека, а также грубых волос, растущих на голове, в подмышечных впадинах, на лобке, а у мужчин еще и на лице. Свободные нервные окончания волосяных фолликулов являются механоре-цепторами и возбуждаются при смещении волос или их подергивании. Другая разновидность свободных нервных окончаний имеется в эпидермисе и в сосочковом слое дермы, большинство из них являются ноцицепторами или терморецепторами, но некоторые принадлежат к механорецепторам, которые специфически реагируют на слабое околопороговое раздражение. Предполагается, что при раздражении этой разновидности рецепторов возникают ощущения щекотки и зуда.

Рис. 17.4. Схема распределения механорецепторов в коже человека. Свободные нервные окончания имеют высокий порог раздражения и слабо реагируют на изменение интенсивности стимула. Быстро адаптирующиеся рецепторы (тельца Пачини, рецепторы волосяных фолликулов) служат датчиками скорости действующих стимулов, а медленно адаптирующиеся рецепторы (диски Меркеля, тельца Руффини) являются датчиками интенсивности действующего раздражителя. Наличие нескольких разновидностей рецепторов позволяет передавать афферентные сигналы о разных свойствах одного и того же раздражителя.
Среди инкапсулированных окончаний различают тельца Пачини, Мейсснера, Руффини, диски Меркеля, тактильные тельца Пинкуса—Игго, колбы Краузе. В зависимости от строения и формы капсулы нервные окончания подвержены наиболее сильному воздействию либо в результате давления действующим перпендикулярно раздражителем, либо вследствие бокового смещения капсулы, которая играет роль механического преобразователя энергии внешних стимулов. Большинство инкапсулированных рецепторов содержится в лишенной волос коже пальцев рук и ног, ладоней и подошв, лица, губ, языка, сосков и половых органов, где они распределены с различной плотностью и на разной глубине. Тельца Пачини имеются также в сухожилиях, связках и брыжейке.
Механорецепторы кожи различаются по скорости адаптации к действующему раздражителю. Быстроадаптирующиеся (фазные) рецепторы возбуждаются только в момент смещения кожи и волос и служат датчиками скорости воздействия стимула. Это свойство присуще тельцам Мейснера, рецепторам волосяных фолликулов и особенно тельцам Пачини, способным реагировать на изменения скорости продолжающего свое действие стимула. Медленно адаптирующиеся (тонические) рецепторы не прекращают генерировать потенциалы действия при продолжительном действии раздражителя, если он оказывает давление на кожу: такие рецепторы служат датчиками интенсивности действующего стимула (тельца Руффини, диски Меркеля).
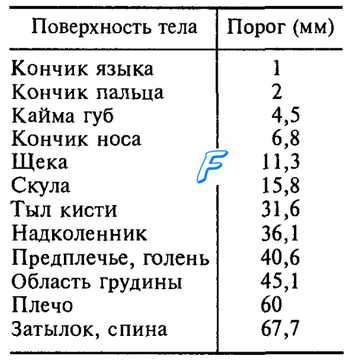
Таблица 17.1. Пространственный двухточечный порог в разных участках тела
Площадь рецептивных полей сенсорных нейронов, иннервирующих тельца Мейснера и диски Меркеля, составляет в среднем около 12 мм2, а у нейронов с окончаниями в виде телец Пачини и Руффини она на порядок больше. Рецептивные поля различающихся своими рецепторами сенсорных нейронов перекрываются, поэтому при действии на кожу комплекса стимулов одновременно возбуждаются разные виды рецепторов, что позволяет ощущать все динамические и статические свойства такого комплекса. Обработка и анализ информации сигналов от различных рецепторов происходит на высших уровнях сенсорной системы, формирующих комплексное восприятие действующих на поверхность тела стимулов. Плотность меха-норецепторов в разных участках кожи не одинакова, чем определяются разные показатели пространственного дифференциального порога, т. е. наименьшего расстояния между двумя точками, раздражение каждой из которых ощущается раздельно (табл. 17.1). Приведенные в таблице данные не следует считать эталоном, поскольку дифференциальная чувствительность различается у разных людей.
Инкапсулированные рецепторы иннервируются миелинизированными волокнами первичных сенсорных нейронов, которые проводят нервные импульсы в ЦНС со скоростью около 30—70 м/с. Немиелинизированные волокна передают потенциалы действия от свободных нервных окончаний со значительно меньшей скоростью — около 1 м/с, поэтому ощущение действующего на них стимула возникает относительно позже. Центральные отростки первичных сенсорных нейронов входят в спинной мозг в составе задних корешков и разделяются в задних рогах спинного мозга на коллатерали. Восходящие коллатерали достигают переключательных ядер заднего столба продолговатого мозга, откуда специфическая информация передается на противоположную сторону мозга по лемнисковому пути, поступает к проекционным ядрам таламуса, а затем в соматосенсорную кору, с участием которой формируется чувство осязания.
Иннервация кожи: нервные окончания, клетки Меркеля, тельца Руффини, Мейснера, Пачини
а) Чувствительные единицы. Любое нервное волокно, разветвляясь, дает начало нервным окончаниям одного вида. Стволовое нервное волокно и его нервные окончания, выполняющие одни и те же физиологические функции, представляют собой чувствительную единицу. В совокупности с исходным униполярным нейроном чувствительная единица аналогична двигательной единице, описанной в отдельной статье на сайте.
Область, раздражение которой приводит к возбуждению чувствительной единицы, называют рецепторным полем. Чем больше размер рецепторного поля, тем меньшей остротой сенсорной чувствительности обладает данная область: например, в верхней части руки рецепторные поля занимают площадь 2 см 2 , в области запястья — 1 см 2 , на подушечках пальцев — 5 мм 2 .
Чувствительные единицы переплетаются между собой, за счет чего становится возможным одновременное восприятие одним участком кожи разных видов чувствительности.
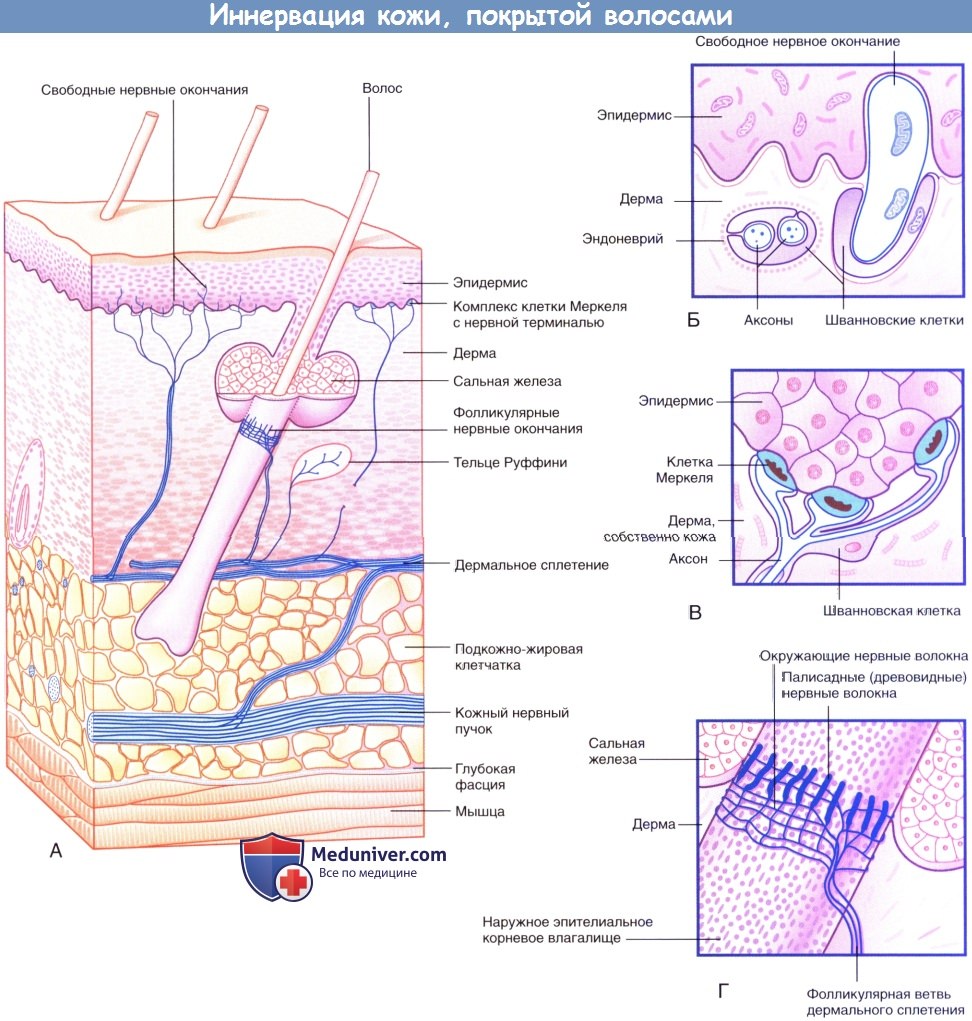
Иннервация кожи, покрытой волосами.
(А) Три морфологических типа чувствительных нервных окончаний в коже, покрытой волосами.
(Б) Свободные нервные окончания в базальном слое эпидермиса.
(В) Комплекс клетки Меркеля с нервной терминалью.
(Г) Палисадные и циркулярные нервные окончания на поверхности наружного корневого влагалища волоса.
б) Нервные окончания:
1. Свободные нервные окончания. По мере приближения к поверхности кожи многие чувствительные нервные волокна утрачивают периневральную, а затем и миелиновую оболочку (в случае ее наличия). Впоследствии нервные волокна разветвляются и формируют субэпидермальное нервное сплетение. Аксон освобождается от оболочек, сформированных шванновскими клетками, что позволяет ему, разветвляясь между коллагеновыми пучками дермы, образовывать дермальные нервные окончания, а внутри эпидермиса — эпидермальные нервные окончания.
Функции. Некоторые чувствительные единицы со свободными нервными окончаниями являются терморецепторами, иннервирующими расположенные на поверхности кожи «тепловые точки» или «холодовые точки». Кроме того, в коже существуют два основных типа ноцицепторов (рецепторов болевой чувствительности), которые также имеют свободные нервные окончания: а-дельта-механоноцицепторы и полимодальные С-ноцицепторы. А-дельта-механоноцицепторы иннервируются тонкими миелинизированными волокнами Аδ-типа и воспринимают существенную механическую деформацию кожи (возникающую, например, при щипке пинцетом). Полимодальные С-ноцицепторы реагируют на болевые стимулы разного вида — механическую деформацию, сильное нагревание или охлаждение (это характерно лишь для некоторых рецепторов), воздействие химических раздражителей. Именно эти рецепторы отвечают за реализацию аксон-рефлекса.
2. Фолликулярные нервные окончания. Нервные окончания волосяного фолликула представлены палисадными нервными волокнами, образованными обнаженными терминалями миелинизированных нервных волокон, расположенными на поверхности наружного корневого влагалища волосяных фолликулов ниже уровня сальных желез, а также циркулярными нервными окончаниями. Каждая фолликулярная единица иннервирует несколько волосяных фолликулов и образует множественные перекресты. Фолликулярные единицы — быстро адаптирующиеся: они возбуждаются при изменении положения волос, однако при сохранении этого положения возбуждения не происходит. Человек, одеваясь, чувствует давление одежды, но затем за счет быстрой адаптации вскоре перестает ощущать ее прикосновение. Иннервация волос у других млекопитающих организована сложнее. Иннервация волосяных фолликулов осуществляется тремя типами механорецепторов, каждый из которых передает информацию определенным структурам мозга, что свидетельствует о важности выполняемой ими чувствительной функции.
3. Комплексы клетки Меркеля с нервной терминалью. Нервная терминаль, расширяясь в области базального слоя эпидермальных гребешков и бороздок, образует комплекс с осязательным тельцем овальной формы — клеткой Меркеля. Комплексы клетки Меркеля с нервной терминалью — медленно адаптирующиеся. В ответ на продолжительное давление (например, при удержании ручки или ношении очков) эти комплексы непрерывно генерируют нервные импульсы. Комплексы клетки Меркеля с нервной терминалью особенно хорошо распознают края удерживаемых в руке предметов.
4. Инкапсулированные нервные окончания. Капсулы описанных ниже свободных нервных окончаний состоят из трех слоев: наружный слой представлен соединительной тканью, средний — периневральным эпителием, а внутренний — видоизмененными шванновскими клетками (телоглией). Инкапсулированные нервные окончания являются механорецепторами, преобразующими механическое воздействие в нервный импульс.
• Тельца Мейснера в большом количестве находятся в подушечках пальцев и расположены вблизи бороздок эпидермиса. Тельца представляют собой клетки овальной формы, внутри которых аксоны располагаются зигзагообразно между уплощенными клетками телоглии. Тельца Мейснера— быстро адаптирующиеся, вместе с медленно адаптирующимися комплексами клетки Меркеля с нервной терминалью они обеспечивают точное восприятие текстур (например, текстуры ткани одежды или поверхности дерева), а также рельефных поверхностей (например, шрифта Брайля). Такие кожные рецепторы способны воспринимать изменение рельефа поверхности даже на высоту 5 нм.
• Тельца Руффини присутствуют как на гладкой коже, лишенной волос, так и на коже с волосами. Они воспринимают плавные скользящие касательные прикосновения и являются медленно адаптирующимися. Внутреннее строение телец сходно со строением сухожильных органов Гольджи: аксоны образуют разветвления в центральной части телец, представленной коллагеновыми волокнами.
• Тельца Пачини по величине соответствуют размерам рисового зерна. В области кисти имеется около 300 телец, которые преимущественно сконцентрированы на боковых участках пальцев и ладони. Тельца Пачини расположены подкожно, близко к надкостнице. Несколько слоев периневрального эпителия внутри соединительнотканной капсулы расположены овально и по форме напоминают луковицу в разрезе. В центральной части тельца Пачини несколько пластинок телоглии окружают единичный аксон, который, попадая в тельце, утрачивает миелиновую оболочку. Тельца Пачини — быстро адаптирующиеся рецепторы преимущественно вибрационной чувствительности. Эти структуры особенно восприимчивы к вибрации костной ткани: большое количество телец расположено в надкостнице длинных трубчатых костей.
Тельца Пачини генерируют один или два нервных импульса при сдавлении и столько же — при прекращении воздействия. В коже ладоней тельца Пачини функционируют по групповому принципу: более 120 телец активируются одновременно, когда человек берет в руку какой-либо предмет (например, апельсин), и когда отпускает его. В связи с этим тельца Пачини считают «детекторами событий» в ходе манипуляций предметами.
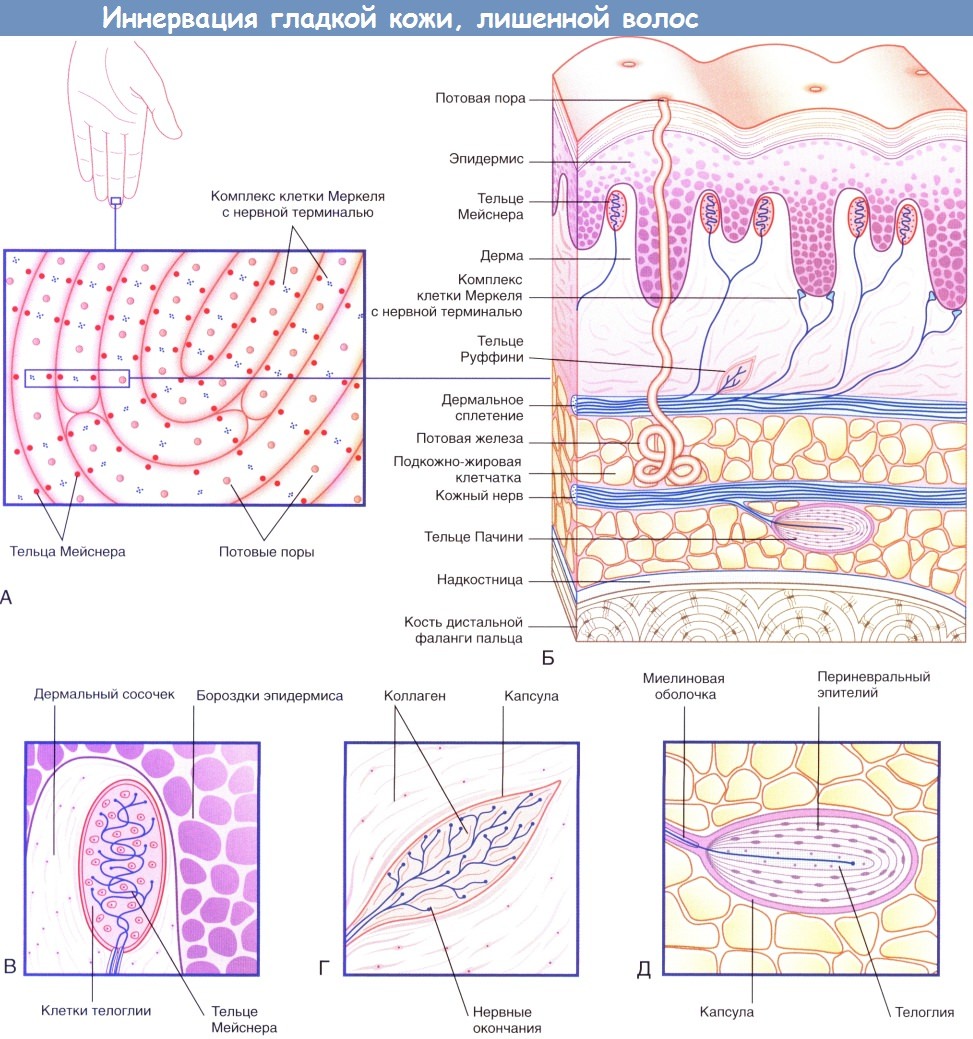
Иннервация гладкой кожи, лишенной волос.
(А) На подушечках пальцев располагаются нервные окончания двух видов.
(Б) На схеме строения участка кожи с изображения (А) представлены четыре типа чувствительных нервных окончаний.
(В) Тельца Мейснера.
(Г) Тельца Руффини.
(Д) Тельца Пачини.
Специалисты по физиологии чувствительности выделяют следующие виды рецепторов, локализованных в коже пальцев.
• Комплексы клетки Меркеля с нервной терминалью — медленно адаптирующиеся рецепторы I типа (MAP I).
• Тельца Мейснера — быстро адаптирующиеся рецепторы I типа (БАР I).
• Тельца Руффини — медленно адаптирующиеся рецепторы II типа (MAP II).
• Тельца Пачини — быстро адаптирующиеся рецепторы II типа (БАР II).
Восприятие ощущений манипуляций с трехмерным предметом вне поля зрения человека в основном обеспечивается за счет мышечных (направляющихся преимущественно от мышечных веретен) и суставных (направляющихся от суставных капсул) афферентных нервных волокон. Кожные, мышечные и суставные афференты независимо друг от друга передают информацию в контралатеральную соматосенсорную зону коры головного мозга. Три различных вида информации объединяются на клеточном уровне в задней части контралатеральной теменной доли, отвечающей за тактильную и визуальную пространственную чувствительность. Тактильную пространственную чувствительность называют стереогнозом. В клинической практике для определения стереогноза пациента просят определить, какой предмет он держит в руках (например, ключ), не смотря на него. Кожные ощущения при периферических нейропатиях описаны в отдельной статье на сайте.
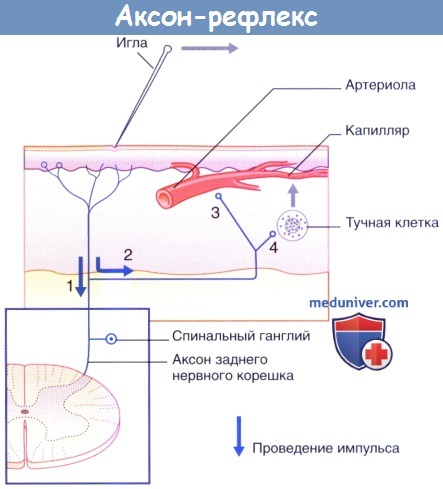
в) Нейрогенное воспаление - аксон-рефлекс. При раздражении чувствительной кожи острым предметом линия контакта практически мгновенно приобретает красный цвет, что обусловлено расширением капилляров в ответ на повреждение кожи. Спустя несколько минут расширение артериол приводит к увеличению зоны гиперемии, а экссудация плазмы из просветов капилляров вызывает формирование бледного отечного валика. Этот феномен представляет собой «тройную реакцию» кожи на раздражение. Формирование зон гиперемии и отечного валика обусловлено аксон-рефлексом чувствительных кожных нервов. Происходящие процессы описаны в соответствии с нумерацией на рисунке ниже.
1. Полимодальные ноцицепторы преобразуют действие болевого раздражителя в нервные импульсы.
2. Аксоны посылают нервные импульсы в центральную нервную систему не только в обычном ортодромном направлении, но и в противоположном антидромном направлении от мест бифуркации к прилежащим участкам кожи. Ответная реакция ноцицептивных нервных окончаний на антидромную стимуляцию проявляется в высвобождении пептидных веществ, среди которых в большом количестве представлена субстанция Р.
3. Субстанция Р связывается с рецепторами на стенках артериол и вызывает их расширение, что приводит к появлению гиперемии.
4. Кроме того, субстанция Р связывается с рецепторами на поверхности тучных клеток, что приводит к высвобождению из них гистамина. Гистамин увеличивает проницаемость капилляров, за счет чего происходит местное накопление тканевой жидкости, обусловливающее возникновение бледного отечного валика.
г) Лепра. Возбудитель лепры — микобактерия, которая проникает в организм человека через мельчайшие повреждения кожи и, распространяясь проксимально по периневрию кожных нервов, вызывает гибель шванновских клеток. Утрата миелиновой оболочки на определенных участках крупных нервных волокон («сегментарная демиелинизация») приводит к нарушению проведения нервных импульсов. Вследствие ответной воспалительной реакции на внедрение возбудителя происходит сдавление всех аксонов, что приводит к валлеровской дегенерации нервов и значительному разрастанию их соединительнотканных оболочек. В результате этого на коже пальцев верхних и нижних конечностей, а также на носу и ушах формируются участки, лишенные чувствительности. Поскольку защитная функция кожной чувствительности нарушается, эти участки становятся более подверженными травматизации, что приводит к повреждению тканей. По мере прогрессирования заболевания возникает двигательный паралич, обусловленный поражением стволов смешанных нервов, расположенных проксимально по отношению к точкам отхождения их кожных ветвей.
д) Резюме. Направляющиеся к коже нервы разветвляются и образуют дермальное нервное сплетение. Чувствительные нервные волокна дермального сплетения разветвляются и перекрывают друг друга. Каждое стволовое нервное волокно и его рецепторы формируют чувствительную единицу. Область, иннервируемую стволовым нервным волокном, называют его рецептивным полем.
К чувствительным единицам со свободными нервными окончаниями относят рецепторы температурной чувствительности, а также механические и температурные рецепторы болевой чувствительности. Рецепторы волосяных фолликулов—быстро адаптирующиеся осязательные механорецепторы, которые активируются только при движении волос. Комплексы клеток Меркеля с нервными терминалями обеспечивают восприятие края предметов, их относят к медленно адаптирующимся.
Инкапсулированные нервные окончания являются механорецепторами. Тельца Мейснера расположены в пространствах между гребешками эпидермиса гладкой кожи, их относят к быстро адаптирующимся. Тельца Руффини—рецепторы растяжения кожи—расположены вблизи ногтей и волосяных фолликулов, их относят к медленно адаптирующимся. Тельца Пачини—подкожные быстро адаптирующиеся нервные окончания, обладающие вибрационной чувствительностью и являющиеся «детекторами событий». На уровне задней части теменной доли коры головного мозга происходит объединение кодированной информации, полученной от кожи, мышц и суставов, что способствует осуществлению тактильного восприятия и стереогностической чувствительности.
Механорецепторы — (греч. mechane – орудие, приспособление, лат. recipio – брать, принимать) рецепторы представляющие собой окончания чувствительных нервных волокон, реагирующие на различные механические стимулы такие как: прикосновение, давление, растяжение, колебания и т.п., действующие извне, или возникающие во внутренних органах организма. В коже они расположены на разной глубине и в различных ее структурных образованиях. Большинство этих рецепторов являются свободными нервными окончаниями чувствительных нервов. Часть их находится в разного рода капсулах.
У человека существуют четыре главных типа таких рецепторов на гладкой коже (то есть коже, лишенной волос): тельце Пачини, сухожильный орган Гольджи .
Механорецепторы, реагирующие на тактильные стимулы (прикосновение к коже или давление) бывают быстро или медленно адаптирующимися.
К быстро адаптирующимся относятся рецепторы волосяных фолликулов волосистой части кожи , тельца Мейснера безволосой (голой) кожи и тельца Пачини подкожной ткани. Рецепторы волосяных фолликулов и тельца Мейснера предпочтительно отвечают на стимулы, поступающие с частотой примерно 30-40 Гц, тогда как тельца Пачини - на стимулы с частотой примерно 250 Гц.
Медленно адаптирующиеся кожные механорецепторы - это диски Меркеля и тельца Руффини. Диски Меркеля имеют точечные рецептивные поля, а тельца Руффини активируются при растяжении кожи, причем даже на некотором расстоянии от рецептора.
У всех этих рецепторов миелинизированные аксоны, большинство из которых принадлежит к группе Aбета, но один класс рецепторов волосяных фолликулов, рецептор опускания волоса, снабжен Aдельта-волокнами.
Содержание
Виды механорецепторов
вестибулярного аппарата), реагируют на ускорения и вибрации при наклоне тела или головы.
Тактильные механорецепторы
Механические стимулы воспринимается рецепторами двух типов:
- Механорецепторами, которые оценивают давление. Они сравнительно медленно адаптируются к внешнему воздействию и обладают плохим временным разрешением.
-
, которые реагируют на возникающую вибрацию. Тельца Пачини воспринимают воздействующую на кожу вибрацию с частотой около нескольких сотен герц. При этом, тельца Пачини работают в комплексе с выростами кожи, которые формируют отпечатки пальцев. Эти регулярные выступы играют роль частотного фильтра и позволяют выделить только полезный сигнал, что позволяет намного точнее определять рельеф ощупываемых предметов. [2]
Барорецепторы
Проприорецепторы
Проприорецепторы - рецепторы мышц (например мышечные веретена и сухожильные рецепторы (органы) Гольджи). Являются как неспецифическими (рецепторы расположены в мышцах и суставах конечностей), так и специфическими (рецепторы расположены в дыхательных мышцах). Импульсация от проприорецепторов распространяется преимущественно к спинальным центрам дыхательных мышц, а также к центрам головного мозга, контролирующим тонус скелетной мускулатуры. Активация проприорецепторов в момент начала физической нагрузки является основной причиной увеличения активности дыхательного центра и повышения вентиляции лёгких. Проприорецепторы межреберных мышц и диафрагмы рефлекторно регулируют ритмическую активность дыхательного центра продолговатого мозга в зависимости от положения грудной клетки в различные фазы дыхательного цикла, а на сегментарном уровне — тонус и силу сокращения дыхательных мышц.
В тканях дыхательных путей расположено два основных типа механорецепторов, импульсы от которых поступают к нейронам дыхательного центра: быстро адаптирующиеся, или ирритантные, рецепторы и рецепторы растяжения.
Читайте также: