
Погруженный эпителий это в зоологии
Обновлено: 26.04.2024
Эпителии кожного типа. Эпидермис. Многослойные эпителии.
Эпителии кожного типа развиваются из кожной эктодермы и прехордальной пластинки. Из кожной эктодермы возникают: многослойный плоский ороговевающий эпителий кожи (эпидермис), многослойный плоский неороговевающий эпителий роговицы, эпителий преддверия ротовой полости, эпителии слюнных, потовых, сальных и молочных желез, переходный эпителий мочевыводящих путей и др.
Из прехордальной пластинки развиваются многослойный плоский неороговевающий эпителий пищевода, многорядный мерцательный эпителий воздухоносных путей, однослойный альвеолярный эпителий легких, эпителий щитовидной, околощитовидной, вилочковой желез и передней доли гипофиза.
По своему строению эпителии кожного типа могут быть многослойные, многорядные и однослойные. Многослойные эпителии состоят из нескольких клеточных слоев, из которых лишь базальный слой прилежит к базальной мембране. Клетки базального слоя — эпителиоциты — способны интенсивно делиться митозом. Они служат источником пополнения клеточного состава вышележащих слоев. Базальные эпителиоциты имеют призматическую форму. По мере смещения этих клеток в поверхностные слои они постепенно уплощаются. В многослойном плоском ороговевающем эпителии поверхностный слой образуют роговые чешуйки.
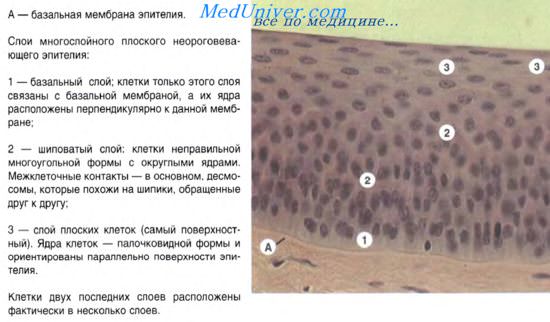
Пограничное положение большинства эпителиев обусловливает определенную цитоархитектонику ткани, а также специфические особенности внутренней структуры клеток и их объединения за счет формирования различных типов межклеточных контактов.
Эпидермис является наиболее типичной разновидностью среди покровных эпителиев. Это полидифферонная ткань. Эпителиальный дифферон развивается из материала кожной эктодермы, отличается стойкой детерминированностью. Диффероны меланоцитов, клеток Лангерганса и клеток Меркеля развиваются из иных источников. Эпителиальный дифферон формирует многослойный пласт ороговевающих клеток (многослойный плоский ороговевающий эпителий). В нем различают слои: базальный, шиповатый, зернистый и роговой. В базальном слое находятся малодифференцированные клетки (базальные эпителиоциты) призматической формы, которые путем митотического деления обеспечивают обновление клеточного состава ткани. После митоза эти клетки перемещаются в вышележащий — шиповатый — слой, образуя клетки многоугольной формы. Клетки шиповатого слоя (шиповатые, крылатые, или остистые, эпителиоциты) имеют в цитоплазме специализированные структуры — тонофиламенты. При световой микроскопии агрегаты тонофиламентов описывают как тонофибриллы. За счет опорных свойств последних достигается механическая прочность клеточного пласта. Между клетками образуются связующие комплексы, или межклеточные контакты — десмосомы.
Следующую стадию дифференцировки составляют уплощенные эпителиоциты зернистого слоя. В цитоплазме этих клеток кроме тонофиламентов синтезируются и накапливаются белки — филаггрин и кератолинин. Ядра зернистых клеток постепенно пикнотизируются, органеллы распадаются под влиянием внутриклеточных ферментов.
Блестящий слой хорошо выявляется только в эпидермисе ладоней и подошв при световой микроскопии. Его образуют плоские постклеточные структуры — кератиноциты, в которых ядра и органеллы исчезают. Из последних образуются роговые чешуйки поверхностного слоя. Они имеют вид 14-гранника. Между чешуйками находится цемонтирующее вещество, богатое липидами (церамиды и др.). Роговые чешуйки имеют плотную оболочку (толщиной 15 им), образованную кератолинином (инволюкрином), ковалентно связанным с оболочкой чешуйки. Содержимое чешуйки заполнено фибриллами зрелого кератина, который характеризуется водонерастворимостью и высокой стойкостью к химическим агентам. Созревание кератина — это агрегация филаментов и обогащение серой за счет образования внутримолекулярных поперечных дисульфидных связей. Этот процесс инициируется филаггрином и происходит при переходе эпителиоцитов из зернистого слоя в роговой. Самые поверхностные слои чешуек постепенно утрачивают связи друг с другом и слущиваются.
Разновидностями многослойных эпителиев являются кубические и призматические эпителии, например, выводных протоков слюнных желез и некоторых других органов, а также многослойный плоский неороговеващий эпителий роговицы. Последний состоит из базального, шиповатого и слоя плоских эпителиоцитов.
Особый вид — переходный эпителий мочевыводящих путей. Он образован базальным, промежуточным и поверхностным слоями. Базальный (камбиальный) слой образован мелкими эпителиоцитами. Полигональные эпителиоциты располагаются в промежуточном слое, а крупные — 2-3-ядерные эпителиоциты — в поверхностном слое. При растяжении мочевого пузыря его стенка уплощается и эпителий растягивается, становится тонким, двуслойным и наоборот, при сокращении — эпителий утолщается. Эпителиоциты промежуточного слоя, не теряя связи с базальной мембраной, становятся грушевидными, а поверхностные — куполообразные.
Многорядный эпителий (ложномногослойный) содержит клетки разной формы. Производными эпителиального дифферона являются реснитчатые, вставочные эпителиоциты, бокаловидные экзокриноциты и эндокриноциты. Располагаются все клетки на базальной мембране. Но вследствие разной высоты ядра эпителиоциов находятся на разных уровнях, что создает впечатление многослойности.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Эта группа, справедливости ради надо отметить, является довольно сборной. Отнесённые сюда эпителии характерны для сравнительно низко организованных животных. Тем не менее, все эти ткани являют примеры чрезвычайно эффективного решения стоящих перед ними функциональных задач, зачастую очень непростых и многообразных.
Архаичные черты строения имеют покровные эпителии губок и некоторых кишечнополостных. Такой эпителий представлен однослойным (в том числе многорядным) пластом плоских, кубических или призматических клеток. Его древность проявляется в слабой детерминированности входящих в состав клеток (например, у гидры этот эпителий может возникать из железистых клеток кишечного эпителия). Кроме того, у кишечнополостных широко распространены эпителиально-мышечные клетки (как следует из названия, сочетающие функции, «впоследствии» выполняемые разными тканями).
Примером однослойных эпителиев, сохранивших, по-видимому, некоторые черты первичных, наиболее древних кожных эпителиев, является погружённый эпителий бескишечных турбеллярий (рис. 3). Характерная его особенность – отсутствие чёткой границы между ним и подлежащей паренхимой. Базальные части клеток, содержащие ядра, свободно располагаются среди клеток паренхимы и как бы погружены в неё (что и обусловило название). Собственно пласт, обеспечивающий выполнение функции этого эпителия, образуют лишь расширенные апикальные части клеток, имеющие межклеточные контакты (в отличие от описанных ранее их нижних частей). На этих апикальных частях развиты многочисленные реснички, которые вместе с мышечными клетками паренхимы образуют локомоторный аппарат этих животных.
Рис. 3. Погружённый и псевдокутикулярный эпителии (по А. А. Заварзину, 2000): а) – погружённый ресничный эпителий бескишечных турбеллярий
(1 – эпителиальные клетки, 2 – паренхимные клетки, 3 – мышечная клетка);
б) – погруженный эпителий цестод (1 – тегумент, псевдо-кутикулярная пластинка, 2 – ядросодержащие части синцития, 3 – базальная мембрана,
4 – продольные и кольцевые мышечные волокна, 5 – паренхимные клетки,
6 – микротрихии).
У остальных турбеллярий (и большинства групп многоклеточных) покровный эпителий чётко обособлен от подлежащих тканей базальной мембраной и представляет собой однослойный (одно- или многорядный) клеточный пласт.
У цестод и трематод - эндопаразитических червей - формируется специализированный псевдокутикулярный эпителий (рис.3). Особенно хорошо его особенности выражены у цестод (лишённых пищеварительной системы). Этот эпидермис эффективно решает две совершенно разные (если не противоположные) функциональные задачи – защиту от действия гидролитических ферментов кишечника хозяина и поглощение находящихся в нём питательных веществ. Это достигается за счёт тегумента (или псевдокутикулярной пластинки). Он образован апикальными частями эпителиальных клеток, которые, соединяясь боковыми поверхностями, формируют синцитий. Тегумент отделён от подлежащей паренхимы базальной мембраной, которая прерывается только в местах его связи с базальными частями клеток, располагающимися в паренхиме; в последних находятся ядра эпителиоцитов (аналогично погружённому эпителию). (Тем псевдокутикула отличается от собственно кутикулы, являющейся неклеточным образованием – см. ниже).
Однослойный эпителий, имеющий синцитиальное строение, характерен и для других групп животных: некоторых турбеллярий, коловраток, скребней, большинства нематод.
Особого внимания заслуживает многорядный мерцательный эпителий, который образует кожные покровы или покрывает отдельные участки поверхности тела очень разнообразных животных (рис. 4). И потому, являясь весьма распространённым с точки зрения систематики, этот эпителий также один из наиболее распространённых и с точки зрения анатомической, так как участвует в образовании весьма разнообразных органов. Причиной этого является наличие на апикальных концах эпителиоцитов ресничек, являющихся универсальным локомоторным (или гидрокинетическим) клеточным органоидом, успешно решавшим (и решающим) задачу движения ещё у одноклеточных (строение ресничек и механизм их движения подробно рассматривается в курсе цитологии).
Кроме того, важной эволюционной новацией этого эпителия явилась дифференциация входящих в его состав клеток на:
1) т. н. мерцательные, снабжённые ресничками, то есть выполняющие основную функцию этого эпителия,
2) камбиальные (т. н. вставочные или клиновидные,
3) секреторные (бокаловидные), обеспечивающие универсальную защиту самого эпителия посредством секреции слизи. Основу последней составляют муцины (мукополисахариды).
Благодаря такой структуре мерцательный эпителий входит в состав тех органов, в которых необходима локомоция – либо самого организма относительно наружной среды, либо жидкостей в его полостях (в половых путях, в дыхательных путях наземных позвоночных и др.). При этом в разных органах в силу специфики выполняемых ими задач в состав этого эпителия могут входить и другие клетки, в том числе других дифферонов.

Рис. 4. Многорядные эпителии (по А. А. Заварзину, 2000): а, б – кожный железистый (а) и ресничный (б) эпителий голожаберных моллюсков;
в – эпителий дыхательных путей млекопитающих; г – покровный эпителий немертин; 1-3 – дифферен-цированные клетки (1 – ресничные; 2 – железистые, 3 – «защитные»), 4 – базальные и 5 – малодифференцированные (вставочные) клетки, 6 – слизь.
Кутикулярные эпителии
Эта группа объединяет эпидермисы, пожалуй, наиболее широко распространённые. Их характерной особенностью является то, что лежащие в один слой эпителиоциты продуцируют поверх своих апикальных концов кутикулу – прочное неклеточное образование. В первую очередь она эффективно осуществляет функцию защиты от разнообразных внешних факторов. За счёт гипертрофии своей прочности кутикулой же у многих животных обеспечивается и опорная функция, формирование так называемого экзоскелета.
Несмотря на большое разнообразие, строение кутикулы всех животных принципиально сходно и построено по принципу «композитных» материалов - то есть кутикулярная пластинка состоит из: а) фибриллярных структур и б) цементирующего аморфного вещества – матрикса. При этом обычно кутикула в той или иной степени находится под контролем эпителиоцитов (чаще посредством их выростов, микроворсинок).
Кроме того, в составе кутикулярных эпителиев нередко имеются и различные секреторные клетки.
Кутикула также может формировать производные, выполняющие дополнительные функции - например, щетинки, которые, по-видимому, в эволюции возникали неоднократно.
На основе структурных различий можно выделить три основные разновидности кутикулярных эпителиев: а) артроподного, б) аннелидного и в) нематодного типов (А. А. Заварзин).
Артроподного типа кутикулярный эпителий(рис. 5).Такие эпителии широко распространены в разных группах животных; наибольшего развития они достигают у членистоногих и оболочников.
Построена такая кутикула на основе фибрилл, образованных линейными молекулами нейтральных полисахаридов (обычно целлюлозы или хитина). Основу матрикса составляют кислые полисахариды (гликозаминогликаны), которые в ряде случаев (например, у наземных членистоногих), замещаются белками и липидами. В большинстве случаев вся кутикулярная пластинка или её наружные слои стабилизируются с помощью ковалентных связей между белковыми молекулами. Это обеспечивается либо за счёт многочисленных дисульфидных связей, или путём так называемого фенольного задубливания, или склеротизации (образования связей между аминогруппами белковых молекул через бензольные кольца хинонов).

Рис. 5. Кутикулярный эпителий членистоногих (по А. А. Заварзину, 2000):
I – гиподерма, II – протокутикула, III – эпикутикула; 1 – эпите-лиальные клетки гиподермы, 2 - эндокутикула, 3 – экзокутикула I, 4 – экзокутикула II,
5 – базальная мембрана, 6 – эноцит, 7 – одноклеточная гиподермальная железа, 8 – цитоплазматические отростки (микровосинки), 9 – протоки гиподермальных желёз, 10 – чувствительные волоски.
Нередко дополнительная механическая прочность кутикулы достигается за счёт биоминерализации – импрегнации кутикулы карбонатами или фосфатами кальция или соединениями кремния, железа, цинка и др., благодаря чему формируются весьма прочные структуры (мандибулы насекомых, радула моллюсков и т. п.).
Особенно сложное строение имеет кутикула наземных членистоногих, включающая в себя несколько слоёв разного строения.
При этом мощная толстая нерастяжимая кутикула в ходе роста животного требует периодической замены посредством линьки – процесса сложного и опасного.
Линька начинается с активизации синтетических процессов в клетках, чему часто предшествует усиленная их пролиферация; увеличение количества клеток приводит к изменению их формы: они становятся кубическими или даже высокопризматическими. Далее клетки апикальными концами выделяют под кутикулярную пластинку гидролитические ферменты, разрушающие отростки клеток и нижнюю часть кутикулы. Образующиеся в результате гидролиза этих компонентов низкомолекулярные продукты всасываются гиподермальными клетками и используются организмом для формирования новых белков и хитина, необходимых для построения новой кутикулярной пластинки. Далее эпителиальные клетки выделяют вещества для формирования новой кутикулы.
Однако этот этап линьки не заканчивается задубливанием – в противном случае она не имела бы смысла, так как образовавшаяся таким путём кутикула не увеличивалась бы в размерах. Склеротизация происходит лишь после сбрасывания прежней кутикулы и после заглатывания животным воды или воздуха, благодаря чему мягкая пока кутикула растягивается.
Аннелидного типа кутикулярный эпителий(рис. 6). Для кутикулы аннелидного типа характерно наличие в ней ряда слоёв коллагеновых волокон. Волокна в пределах одного слоя лежат параллельно друг другу, но относительно волокон соседнего слоя – перпендикулярно. Коллагеновые фибриллы этих волокон не имеют поперечной исчерченности (в отличие от коллагена соединительных тканей – см. соответствующий раздел). Над слоями коллагеновых волокон и в промежутках между ними располагаются аморфный и фиброзный матриксы, образованные мукополисахаридами и неколлагеновыми белками.

Рис. 6. Кутикулярный эпителий аннелидного типа (на примере аннелид и погонофор) (по Заварзину А. А., 2000). 1- эпителиальные клетки, 2 – микроворсинки эпителиальных клеток, 3 – коллагеновые волокна, 4 – гликопротеидный матрикс, 5 – наружный слой кутикулы, 6 – сферические тельца, 7 – гликокаликс.
У аннелид кутикула пронизана длинными микроворсинками, выходящими на поверхность. Последние позволяют поглощать органические вещества из окружающей среды. (Существует мнение, что такая кутикула могла возникнуть на основе гипертрофии гликокаликса на поверхности микроворсинок эпителиоцитов).
Несмотря на огромное многообразие червей этой группы, строение их кутикулы принципиально сходно. Подобное строение также имеют кутикулы погонофор, сипункулид и др.
Нематодного типа кутикулярный эпителий (рис. 7). Кутикула нематод может рассматриваться как вариант кутикулы аннелидного типа. Однако этот эпителий – одна из наиболее сложных покровных тканевых систем беспозвоночных. Сложность обусловлена спецификой биологии нематод – эндопаразитической группы – одной из наиболее многочисленных среди современных животных. Их эпидермис эффективно решает две ведущие функциональные задачи – барьерную и опорную.

Рис. 7. Кутикулярный эпителий нематод (по А. А. Заварзину, 2000):
а – принципиальная схема организации, б - кутикулярный эпителий аскариды. 1 – цитоплазма гиподермы, 2 – ядросодержащая часть, 3 – базальная мембрана, 4 – базальный слой кутикулы, 5 – промежуточный слой, 6 – кортикальный слой, 7 – эпикутикула, 8 – полость в промежуточном слое.
Структурно это обеспечивается следующим образом. Гиподерма у большинства нематод имеет синцитиальное строение. Основу кутикулы составляют коллагеновые волокна, имеющие сходное строение с волокнами кутикулы аннелидного типа. Они образуют два нижних слоя этой кутикулы – базальный и промежуточный. Эти слои отличаются «упаковкой» волокон: в базальном слое они объединены в пластинки и ориентированы таким образом, что в соседних пластинках лежат относительно друг друга под углом примерно 135 градусов; в промежуточном слое волокна не имеют строгой ориентации.
Верхний слой – кортикальный - представлен аморфным веществом, образованным белком кутикулином с большим числом ковалентных связей между молекулами. Наконец, на поверхности имеется тонкая эпикутикула, состоящая из белков и липидов.
Столь прочная структура требует периодической линьки, которая и происходит в ходе развития нематод обычно 4 раза.
Однако такая сложная конструкция кутикулы позволяет и модифицировать её в широком диапазоне: у некоторых нематод она может подвергаться фенольному задубливанию, превращая стенку тела в прочную цисту; у других же – практически редуцируется, благодаря чему животное может питаться всасыванием через микроворсинки.
Многослойные эпидермисы
Многослойные эпидермисы характерны для позвоночных животных, что резко отличает их гистологически от беспозвоночных (у которых эпидермис, как видно из предыдущего очерка, однослойный).
У низших позвоночных покровный эпителий имеет небольшое число слоёв, что демонстрирует их происхождение от беспозвоночных.
Наиболее сложно устроен эпидермис наземных позвоночных (Приложение, рис. 5). Это обусловлено особенностями воздушной среды обитания – её нестабильностью, а также потребностью защиты от потерь воды (дегидратации).* Для решения отмеченных функциональных задач на поверхности эпителия Amniota формируется толстый защитный слой, образованный роговыми чешуями – постклеточными структурами, образующимися в результате довольно сложного преобразования исходных клеток. Этим обусловлено наличие в данном эпителии нескольких слоёв клеток. Рассмотрим это строение на примере кожи человека.
1). Непосредственно на базальной пластинке лежит один слой призматических клеток, являющихся стволовыми.
2). Выше располагаются 7-12 слоёв т. н. шиповатых или крылатых клеток (своё название они получили за наличие отростков –
*Кутикулярный эпителий наземных членистоногих и многослойный ороговевающий эпителий наземных позвоночных – яркий пример тканевой конвергенции, вызванной сходством «экстремальных» условий существования.
«шипов» - для плотного примыкания этих клеток друг к другу).
3). Далее располагаются 2-5 рядов слегка уплощённых т. н. зернистых клеток. Своё название они получили за то, что содержат кератогиалиновые зёрна (гранулы), содержащие белки, богатые гистидином и цистеином, а также мукополисахариды; наличие этих гранул является внешним проявлением развивающегося процесса ороговения (кератинизации).
4). Ещё выше располагается блестящий слой. Образующие его клетки утратили ядра и органоиды и заполнены волокнистой массой - элеидином, обладающим двойным лучепреломлением, чем обусловлена своеобразная окраска этого слоя.
5). Наконец, располагающийся на поверхности роговой слой образован уже отмечавшимися роговыми чешуями (корнеоцитами), заполненными воздухом и содержащими белок кератин; межклеточные пространства заполнены нейтральными липидами, способствующими гидроизоляции.
Подобного рода структура эпидермиса позволяет ему, помимо прочего, формировать разнообразные производные (кожные чешуи и щитки, перья, волосы, вибриссы, когти, ногти, рога полорогих животных, копыта и т. п.), которые выполняют у наземных позвоночных различные функции - защитные, опорные, механические, теплоизолирующие и другие.
Мощный роговой слой и его производные у наземных позвоночных так же, как и кутикулярный эпителий, требует замены – линьки: либо единовременной (рептилии), либо постепенной (птицы и млекопитающие).
Вышеперечисленные типы клеток составляют единый эпидермальный дифферон ороговевающих клеток. Помимо него, в эпидермисе человека представлен ряд других дифферонов:
1) меланобласты и меланоциты (синтезируют и содержат пигмент меланин);
2) клетки Лангерганса (их источник – стволовые клетки крови);
3) клетки Меркеля (вырабатывают после редукции тимуса гормоны, подобные синтезируемым им тималину и тимулину; кроме того, к этим клеткам подходят нервные окончания);
4) Т-лимфоциты – (иммунокомпетентные клетки).
Сходно с ороговевающим устроен у позвоночных эпителий, покрывающий роговицу глаза и образующий выстилку полости рта, глотки, пищевода. Но в нём представлены лишь три отмеченных выше слоя – базальный, шиповатый и роговой; клетки верхних слоёв отмирают в нём без ороговения. (Правда, при некоторых нарушениях и в этом эпителии может происходить кератинизация – так, например, на роговице образуется т. н. бельмо).
«Барьер» между средой и организмом. Все границы. Из всех 3х зародышевых листков.
Особенности, обусловленные их предназначением (связанные с барьерной функцией):
-объединение эпителиальных клеток в непрерывные пласты или тяжи;
-отсутствие межклеточного вещества (совсем нет);
- полярность клеток; наличие многочисленных и разнообразных клеточных контактов (в апикальной части – замыкающие пластинки, снизу – десмосомы, полудесмосомы);
- высокая способность к регенерации;
Функции эпителиальных тканей: разграничительная, всасывающая (пищеварительная), железистая, выделительная, осморегулирующая, транспортная (газов), локомоторная.
-по слойности (по количеству слоёв клеток, соприкасающихся с мембраной) – однослойные (все клетки контактируют с базальной мембраной), многослойные, переходный (выстилает органы, подверженные сильному растяжению);
- по рядности (количеству рядов ядер в однослойном эпителии) – однорядные, многорядные;
- по форме клеток – кубический, цилиндрический, плоский (ядро подчиняется форме);

- по степени ороговения (?).
Типы эпителия, их особенности и представительство.
На ранних этапах эволюции эпителий выполняет все функции. Нет параллелизма, потому что у всех групп формировался в разных средах/условиях.
Покровный эпителий.
Впервые у плоских червей (турбелярий). (У кишечнополостных эпителиально-мускульная клетка. Двигает не только клетку, но и тело. Не только барьер, но и мышца – поэтому не эпителий.)

У бескишечных турбелярий эпителий погружённого типа. Погружённый, так как нет базальной мембраны, базальная часть погружена в паренхиму. Нет плотных контактов, реснички для движения. В апикальной части микрафиламенты.

У остальных турбелярий – истинный эпителий нимертинового типа. Однослойный, многорядный, мерцательный с базальной мембраной, комбиальными элементами. У нимертин реснички не выполняют функций движения.
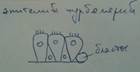
Класс трематоды (сосальщики). Живут в протоках, в агрессивной среде. Верхняя часть эпителия сливается в синцитий (частично или полностью утрачивает границы).
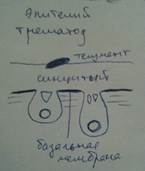
Класс цестоды (ленточные черви). В тонком кишечнике. В синцитие образуются микроворсинки.
У беспозвоночных за счёт кутикулы защита от внешней среды.

Класс нематоды. В почвах. Проблемы с механическим повреждением и высушиванием. Кутикула из 6 слоёв – осн. белок (похож на коллаген), образующий сеть и липидный слой от высыхания. Эпителий сливается в синцитий, потом исчезает, потому что не регенерации.
Класс кольчатые черви. У олигохет (донные пресноводные) микроворсинки, синтезирующие коллагеноподобный туницин. Бляшки не закрываются кутикулой, так как через покровы идёт газообмен.

Ракообразные. Хитинизированная кутикула. хитин способен затвердевать за счёт минерализации (карбонат/фосфат кальция).

Насекомые. Проблемы насекомых – тяжесть, высушивание. Липидная эпикутикула. Не минерализуется, а дубится. При небольшой толщине – высокая прочность.

Хордовые. Увеличение слойности.

Амфибии. Ороговение (кератинизация) – накопление керато-геолинов? – гидрофобные свойства препятствуют испарению. Частичное ороговение – все клетки эпителия живые, так как присутствует кожное дыхание. Полное ороговение - у всех групп, вышедших на сушу. Верхний слой – чешуйки с кератином, мёртвые. Кератинизация мягкая (чешуйки рыхло, связаны не плотно, постоянно слущиваются, хуже защищает от испарения) и твёрдая (у рептилий; ногти; над клювом. Ограничивает рост).
Кишечный эпителий.
На поверхности гликокаликс – слой из гликозаминогликанов, кот. присоединяются к мембране микроворсинок («кисель»). Пристеночное пищеварение. Гладкий ЭПР, митохондрии. Провести расщеплённые вещества в соседнюю ткань. –слизистые клетки в составе этого эпителия, - эндокриноциты – регулируют активность транспорта или активность клеток. Однорядный. Все клетки в одном функциональном состоянии.
Осморегуляторный и выделительный эпителий.
Фильтрационный – из внутрынней среды выкачивает воду с продуктами обмена.
Реабсорбционный – обратно в кровь, что нужно (кубический эпителий, крупное ядро, много микроворсинок, пиноцитозные пузырьки, базальная исчерченность). Отдел похож у всех, различна лишь длина.
Накопительный – есть не всегда (переходный тип эпителия, у кроющих клеток чередуются плотные и тонкие участки, чтобы изменять форму; многоядерный, так как высокий обмен из-за агрессивной среды).

Эпителий со спец. свойствами.
Мезотелий – однослойный, плоский, мезодерм. происх., выстилает полость брюшины, предотвращает спайки между органами, регулирует химизм полостных жидкостей.
Выстилка трахеи – односл., многоряд., мерцател., цилиндрич.
Эндотелий – самый плоский, выстилает все участки кровеносной и лимфатической систем.
Железистый эпителий.
Железы экзокринные ( секрет, через протоки/на границу раздела сред) и эндокринные (гормоны, нет протоков, в кровь или внутр. среду).
Экзокринные. Секреторный отдел – секреторные клетки (,миоэпителиальные клетки).+ Выводящий проток. = паренхима железы. Строма – всё, что окружает и поддерживает паренхиму.
Классификация: по форме секреторного отдела альвеолярные, трубчатые и альвеолярно-трубчатые;
ветвление секреторного отдела – разветвлённое, не разветвлённое;

ветвление протока – сложное и простое;

способ секреции – эккриновый (экзоцитоз), апокриновый,голокриновый;

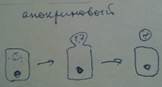

по химизму секрета – слизистые, белковые, смешанные, водно-солевые, сальные.
Как следует из названия, этот эпителий формирует кожные покровы животных.
Уже отмечалось, что это одни из наиболее древних эпителиев (да и тканей вообще), который представлен у всех многоклеточных животных.
В соответствии с изложенной выше гипотезой, функции этого эпителия обусловлены интенсивным контактом с внешней средой. На ранних этапах эволюции его а) барьерная роль сочеталась с б) поглощением питательных веществ из внешней среды и в) выделением в неё вредных продуктов метаболизма, а также г) локомоторной функцией и д) восприятием раздражений из внешней среды.
Позднее для осуществления этих функций стали появляться иные, специальные ткани, а кожный эпителий начал специализироваться на создании совершенного барьера, изолирующего организм от внешней среды, а так же связывющего с ней. Особенно большое значение барьерная функция этих тканей приобретает при выходе животных на сушу; кроме того, у многих животных кожный эпителий берёт на себя и скелетную (опорную) функцию (что будет рассмотрено ниже).
Так как взаимодействие с внешней средой разнообразно, а нередко и противоречиво, а также в силу огромного многообразия условий внешней среды строение эпидермисов весьма варьирует.
Большое многообразие строения кожных эпителиев можно условно разделить на три группы (по А. А. Заварзину), а именно различать эпидермисы:
погружённые, однослойные, многорядные;
кутикулярные;
многослойные.
Такая последовательность в известной степени отображает и последовательное возникновение различных эпидермисов в филогенезе.
2.1.2.1.2. Погружённые, однослойные, многорядные эпидермисы
Эта группа, справедливости ради надо отметить, является довольно сборной. Отнесённые сюда эпителии характерны для сравнительно низко организованных животных. Тем не менее, все эти ткани являют примеры чрезвычайно эффективного решения стоящих перед ними функциональных задач, зачастую очень непростых и многообразных.
Архаичные черты строения имеют покровные эпителии губок и некоторых кишечнополостных. Такой эпителий представлен однослойным (в том числе многорядным) пластом плоских, кубических или призматических клеток. Его древность проявляется в слабой детерминированности входящих в состав клеток (например, у гидры этот эпителий может возникать из железистых клеток кишечного эпителия). Кроме того, у кишечнополостных широко распространены эпителиально-мышечные клетки (как следует из названия, сочетающие функции, «впоследствии» выполняемые разными тканями).
Примером однослойных эпителиев, сохранивших, по-видимому, некоторые черты первичных, наиболее древних кожных эпителиев, является погружённый эпителий бескишечных турбеллярий (рис. 3). Характерная его особенность – отсутствие чёткой границы между ним и подлежащей паренхимой. Базальные части клеток, содержащие ядра, свободно располагаются среди клеток паренхимы и как бы погружены в неё (что и обусловило название). Собственно пласт, обеспечивающий выполнение функции этого эпителия, образуют лишь расширенные апикальные части клеток, имеющие межклеточные контакты (в отличие от описанных ранее их нижних частей). На этих апикальных частях развиты многочисленные реснички, которые вместе с мышечными клетками паренхимы образуют локомоторный аппарат этих животных.

Рис. 3. Погружённый и псевдокутикулярный эпителии (по А. А. Заварзину, 2000): а) – погружённый ресничный эпителий бескишечных турбеллярий (1 – эпителиальные клетки, 2 – паренхимные клетки, 3 – мышечная клетка); б) – погруженный эпителий цестод (1 – тегумент, псевдо-кутикулярная пластинка, 2 – ядросодержащие части синцития, 3 – базальная мембрана, 4 – продольные и кольцевые мышечные волокна, 5 – паренхимные клетки, 6 – микротрихии).
У остальных турбеллярий (и большинства групп многоклеточных) покровный эпителий чётко обособлен от подлежащих тканей базальной мембраной и представляет собой однослойный (одно- или многорядный) клеточный пласт.
У цестод и трематод - эндопаразитических червей - формируется специализированный псевдокутикулярный эпителий (рис.3). Особенно хорошо его особенности выражены у цестод (лишённых пищеварительной системы). Этот эпидермис эффективно решает две совершенно разные (если не противоположные) функциональные задачи – защиту от действия гидролитических ферментов кишечника хозяина и поглощение находящихся в нём питательных веществ. Это достигается за счёт тегумента (или псевдокутикулярной пластинки). Он образован апикальными частями эпителиальных клеток, которые, соединяясь боковыми поверхностями, формируют синцитий. Тегумент отделён от подлежащей паренхимы базальной мембраной, которая прерывается только в местах его связи с базальными частями клеток, располагающимися в паренхиме; в последних находятся ядра эпителиоцитов (аналогично погружённому эпителию). (Тем псевдокутикула отличается от собственно кутикулы, являющейся неклеточным образованием – см. ниже).
Однослойный эпителий, имеющий синцитиальное строение, характерен и для других групп животных: некоторых турбеллярий, коловраток, скребней, большинства нематод.
Особого внимания заслуживает многорядный мерцательный эпителий, который образует кожные покровы или покрывает отдельные участки поверхности тела очень разнообразных животных (рис. 4). И потому, являясь весьма распространённым с точки зрения систематики, этот эпителий также один из наиболее распространённых и с точки зрения анатомической, так как участвует в образовании весьма разнообразных органов. Причиной этого является наличие на апикальных концах эпителиоцитов ресничек, являющихся универсальным локомоторным (или гидрокинетическим) клеточным органоидом, успешно решавшим (и решающим) задачу движения ещё у одноклеточных (строение ресничек и механизм их движения подробно рассматривается в курсе цитологии).
Кроме того, важной эволюционной новацией этого эпителия явилась дифференциация входящих в его состав клеток на:
1) т. н. мерцательные, снабжённые ресничками, то есть выполняющие основную функцию этого эпителия,
2) камбиальные (т. н. вставочные или клиновидные,
3) секреторные (бокаловидные), обеспечивающие универсальную защиту самого эпителия посредством секреции слизи. Основу последней составляют муцины (мукополисахариды).
Благодаря такой структуре мерцательный эпителий входит в состав тех органов, в которых необходима локомоция – либо самого организма относительно наружной среды, либо жидкостей в его полостях (в половых путях, в дыхательных путях наземных позвоночных и др.). При этом в разных органах в силу специфики выполняемых ими задач в состав этого эпителия могут входить и другие клетки, в том числе других дифферонов.
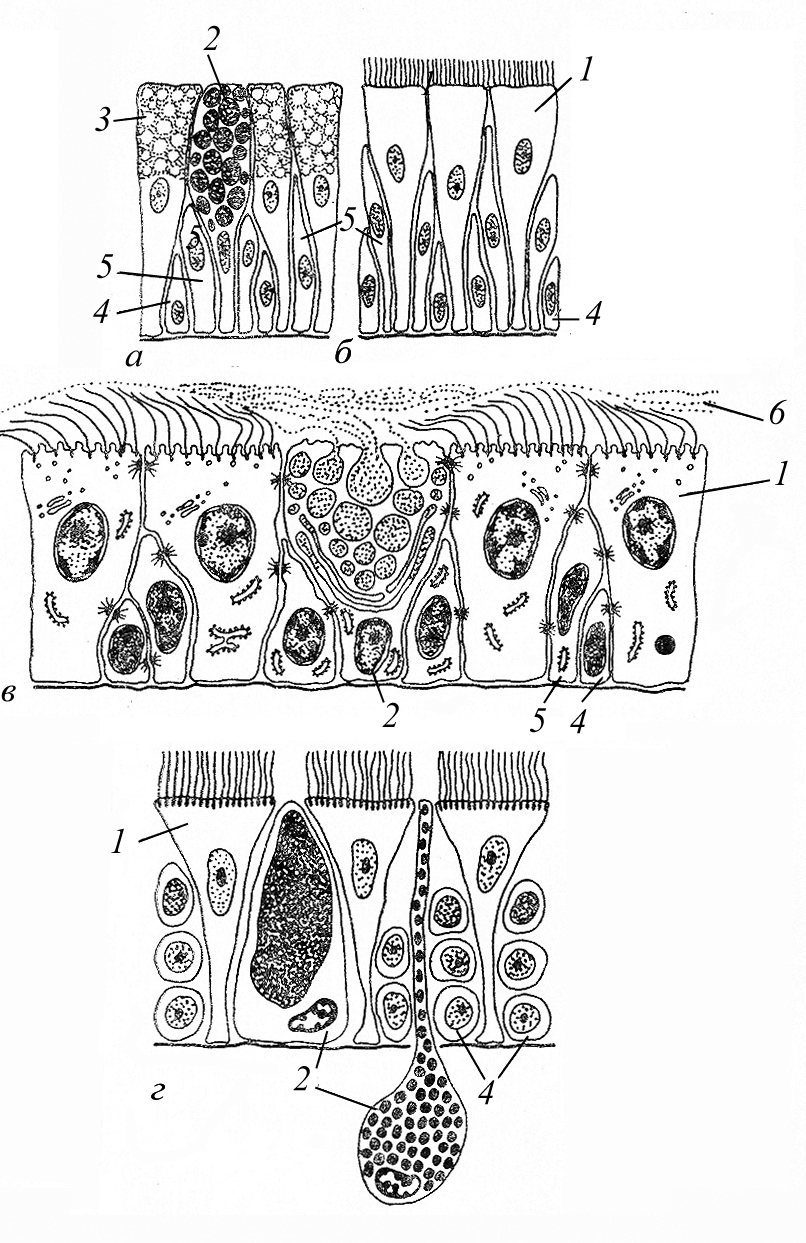
Рис. 4. Многорядные эпителии (по А. А. Заварзину, 2000): а, б – кожный железистый (а) и ресничный (б) эпителий голожаберных моллюсков; в – эпителий дыхательных путей млекопитающих; г – покровный эпителий немертин; 1-3 – дифферен-цированные клетки (1 – ресничные; 2 – железистые, 3 – «защитные»), 4 – базальные и 5 – малодифференцированные (вставочные) клетки, 6 – слизь.
Эпителиальная ткань животных. Простые эпителии.
Эпителиальная ткань животных образует однослойные или многослойные пласты, покрывающие внутренние и наружные поверхности любого организма.
Эпителиальные клетки соединены друг с другом небольшим количеством цементирующего вещества, состоящего в основном из углеводов, и специальными связками — межклеточными контактами. Эпителий подстилает базальная мембрана, состоящая из переплетающихся коллагеновых волокон, заключенных в матрикс. Термин мембрана не следует путать с клеточными мембранами, о которых мы говорили в гл. 5; здесь он означает просто тонкий слой. Матрикс не препятствует диффузии. Поскольку эпителиальные клетки не снабжаются кровеносными сосудами, кислород и питательные вещества поступают к ним путем диффузии из лимфатических сосудов, расположенных в межклеточных пространствах. В эпителий могут проникать нервные окончания.
Функция эпителиальной ткани заключается в защите нижележащих структур от механических повреждений и от инфекции. При постоянных механических воздействиях эта ткань утолщается и кератин изируется, а в тех участках, где клетки слущиваются вследствие постоянного давления или трения, клеточное деление происходит с очень высокой скоростью, так что утраченные клетки быстро замещаются. Свободная поверхность эпителия часто бывает высоко дифференцированной и выполняет функции всасывания, секреторные или экскреторные или же содержит сенсорные клетки и нервные окончания, специализированные к восприятию раздражений.
Эпителиальная ткань делится на несколько типов в зависимости от числа клеточных слоев и от формы отдельных клеток. Во многих частях организма клетки разных типов перемешаны друг с другом, и тогда эпителиальную ткань бывает трудно отнести к какому-либо определенному типу.
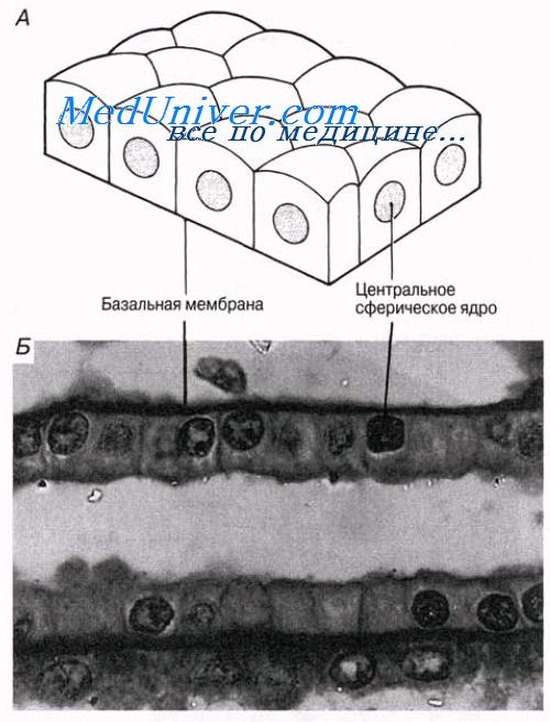
Простые эпителии
Плоский эпителий
Клетки плоского эпителия тонкие и уплощенные. Уплощены они настолько, что ядро образует выпуклость. Края клеток неровные.
Как это ясно видно на изображении поверхности клетки. Соседние клетки плотно соединены друг с другом особыми контактами. Плоский эпителий имеется в боуменовых капсулах почек, в выстилке альвеол легких и в стенках капилляров, где благодаря своей тонкости он допускает диффузию различных веществ. Образует также выстилку полых структур, таких как кровеносные сосуды и камеры сердца, где уменьшает трение при протекании жидкостей.
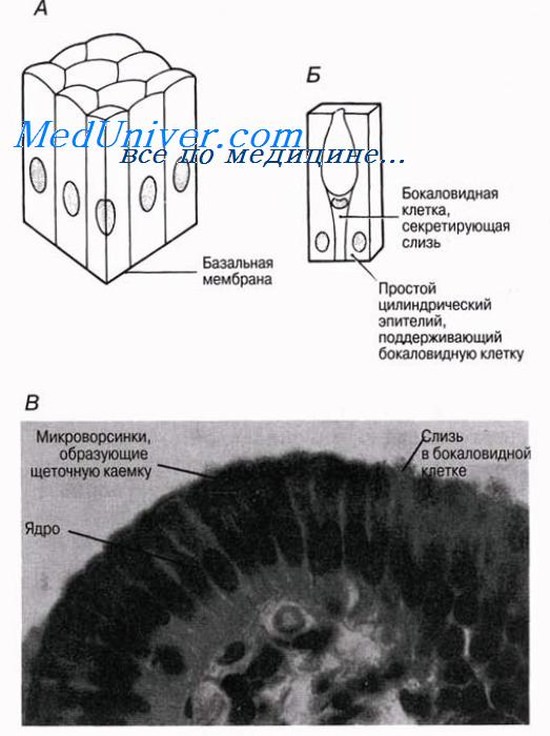
Кубический эпителий
Это наименее специализированный из всех эпителиев. Его клетки имеют кубическую форму и содержат расположенное в центре сферическое ядро. Если смотреть на эти клетки сверху, то видно, что они имеют пяти- или шестиугольные очертания. Кубический эпителий выстилает протоки многих желез, например слюнных желез и поджелудочной железы, а также проксимальные и дистальные почечные канальцы и собирательные трубочки почки в участках, где они не являются секреторными.
Кубический эпителий содержится также во многих железах — слюнных, слизистых, потовых, щитовидной, — где он выполняет секреторные функции.
Цилиндрический эпителий
Клетки этого эпителия высокие и довольно узкие; благодаря такой форме на единицу площади эпителия приходится больше цитоплазмы. В каждой клетке имеется ядро, расположенное на ее базальном конце. Среди эпителиальных клеток часто бывают разбросаны бокаловидные клетки; по своим функциям цилиндрический эпителий может быть секреторным и(или) всасывающим. Нередко на свободной поверхности каждой клетки имеется хорошо выраженная щеточная каемка, образуемая микроворсинками, которые увеличивают всасывающую и секреторную поверхность клетки. Цилиндрический эпителий выстилает желудок; слизь, выделяемая бокаловидными клетками, защищает слизистую желудка от воздействия кислого содержимого и от переваривания ферментами. Он выстилает также кишечник, где опять-таки слизь защищает стенки кишечника от самопереваривания и одновременно создает смазку, облегчающую прохождение пищи. В тонком кишечнике переваренная пиша всасывается через этот эпителий в кровяное русло. Цилиндрический эпителий выстилает и защищает многие почечные канальцы; он имеется также в щитовидной железе и желчном пузыре.
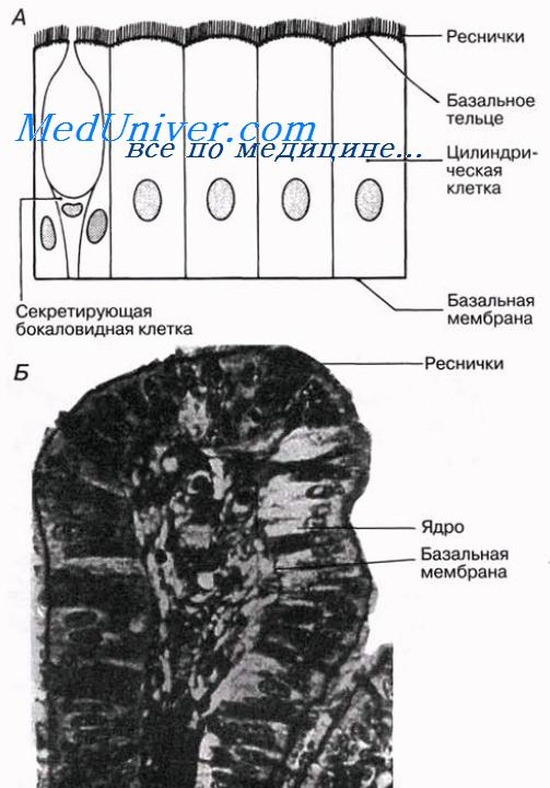
Мерцательный эпителий
Клетки этого эпителия обычно имеют цилиндрическую форму, но несут на своих свободных поверхностях многочисленные реснички. Они всегда ассоциированы с бокаловидными клетками, секретирующими слизь, перетекающую благодаря биению ресничек. Мерцательный эпителий выстилает изнутри яйцеводы, желудочки головного мозга, спинномозговой канал и дыхательные пути (трахеи, бронхи и бронхиолы), обеспечивая перемещение по ним различных веществ. Так, например, вдыхательных путях реснички перемещают слизь вверх, в горло, что облегчает заглатывание твердой пищи. Слизь задерживает бактерии, пыль и другие мелкие частицы, препятствуя их попаданию в легкие.
Псевдомногослойный (многорядный) эпителий
При рассмотрении гистологических срезов этого эпителия создается впечатление, что клеточные ядра лежат на разных уровнях, потому что не все клетки доходят до свободной поверхности ткани. Тем не менее этот эпителий состоит только из одного слоя клеток, каждая из которых прикреплена к базальной мембране. Псевдомногослойный эпителий выстилает мочевые пути и дыхательные пути (трахею, бронхи, бронхиолы, где он покрыт ресничками и состоит из цилиндрических клеток).
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: