
Какой белок содержится в эпителиальных клетках хрусталика
Обновлено: 29.04.2024
Хрусталик. Стекловидное тело. Сосудистая оболочка.
Хрусталик развивается из материала эктодермальной хрусталиковой плакоды, преобразующейся под влиянием глазного бокала в хрусталиковый пузырек. На 5-й неделе эмбриогенеза хрусталиковый пузырек отшнуровывается от эктодермы.
Передняя стенка хрусталикового пузырька состоит из однослойного кубического эпителия, заднюю стенку образуют удлиненные клетки — хрусталиковые волокна. По мере их роста полость пузырька исчезает. В центре развивающегося хрусталика из первичных хрусталиковых волокон формируется ядро хрусталика.
При этом хрусталиковые волокна теряют ядра, а процесс физиологической регенерации белков осуществляется на долгоживущих РНК.
Дальнейший рост хрусталика связан с образованием вторичных хрусталиковых волокон за счет пролиферации клеток, находящихся в экваториальной области. Цитоплазма хрусталиковых волокон содержит прозрачное вещество — белок кристаллин. Внутри хрусталика сосудов и нервов нет. Хрусталик покрыт прозрачной капсулой. На передней поверхности хрусталика под капсулой сохраняется однослойный эпителий, называемый эпителием передней сумки.
Хрусталик поддерживается в глазу в определенном положении посредством ресничного пояска, состоящего из нитей цинновой связки. Нити прикрепляются с одной стороны к ресничному телу, а с другой — к капсуле хрусталика в области его экватора. Изменение степени натяжения цинновой связки сопровождается изменением кривизны поверхностей хрусталика, имеющего форму двояковыпуклой линзы. При этом меняется преломляющая сила хрусталика, благодаря чему возможна аккомодация глаза, т. е. способность четкого видения различно удаленных предметов.
Стекловидное тело.
В эмбриогенезе различают три стадии формирования стекловидного тела: первичное стекловидное тело, состоящее из мезенхимных клеток, проникающих в полость глазного бокала, которые дифференцируются в гиалоциты; вторичное стекловидное тело, для которого характерны редукция сосудов и продукция прозрачного вещества нейроглиальным эпителием внутренней оболочки глаза; и сформированное стекловидное тело.
Оно представляет собой желеобразное прозрачное вещество, содержащее около 99% воды и плотный остов из белка витреина и гиалуроновой кислоты. Стекловидное тело представляет собой основную светопреломляющую среду глаза. Вместе с тем это и своего рода амортизатор, способствующий созданию внутриглазного давления. Стекловидное тело является метаболически активным веществом, участвующим в трофических процессах сетчатки.
В стекловидном теле отсутствуют нервы и сосуды. При электронной микроскопии установлено наличие в нем фибриллярных структур, образующих нитчатый остов, гиалоцитов, макрофагов и лимфоцитов.
Сосудистая оболочка.
Питание глаза осуществляет средняя — сосудистая — оболочка. В сосудистой оболочке различают надсосудистую, сосудистую, сосудисто-капиллярную пластинки, которые состоят из рыхлой волокнистой соединительной ткани с большим количеством меланоцитов. Сосудистая и сосудисто-капиллярная пластинки содержат артерии, вены и капиллярные сети. Здесь происходит переход крови из артериального в венозное русло.
На границе между сосудистой оболочкой и пигментным слоем сетчатки находится мембрана Бруха, представляющая собой тонкий (1-4 мкм) слой коллагеновых и эластических волокон. Через нее происходит диффузное питание сетчатки.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Вода и неорганические соединения. В составе нормального хрусталика среди веществ низкой молекулярной массы 60-65 % занимает вода. Распределение ее в хрусталике неравномерно. Наибольшее количество воды содержится к коре хрусталика, где происходит ее интенсивный обмен с полным замещением за 1–2 часа. Меньше воды в ядре хрусталика, и ее полное замещение происходит здесь в течение 5-20 часов. Вода в хрусталике играет важную роль в состоянии его оптических свойств (светорассеивание и рефракция). С возрастом содержание воды в хрусталике понижается. Изменяется количество воды в хрусталике при развитии различных катаракт как в сторону увеличения, так и уменьшения.
В частности, при старческой катаракте способность белков помутневшего хрусталика связывать воду уменьшается по сравнению с его нормальным состоянием, при этом уровень свободной воды увеличивается.
В составе неорганических соединений хрусталика, составляющих 5–7% зольного остатка, преобладающим катионом является калий, необходимый для реакций гликолиза, синтеза белка на рибосомах, регуляции осмотического давления. Другим активным катионом, регулирующим осмотическое давление в структурах хрусталика, является натрий, располагающийся преимущественно в межклеточных пространствах. При нарушениях нормального соотношения в хрусталике ионов калия и натрия обычно наступает его помутнение. Дисбаланс ионов калия и натрия занимает важное место в развитии старческой и рентгеновской катаракт. В помутневшем хрусталике, как правило, больше натрия и меньше калия, чем в прозрачном.
Ионы магния в хрусталике необходимы для образования комплекса с АТФ, из которого извлекается энергия для процессов активного транспорта, необходимых для метаболических функций питательных веществ.
Ионы кальция оказывают регулирующее влияние на концентрацию других ионов в хрусталике и необходимы для образования высокомолекулярных агрегатов его белков и поддержания нормальной проницаемости мембран.
Фосфор содержится в органических и неорганических фосфатах хрусталики. в частности в АТФ, и на долю их приходится до 39 % зольного остатка хрусталика.
Сера входит в состав сульфатов, цистеина, цистина, метионина и глутатиона хрусталика.
Хлор в составе хлоридов регулирует осмотическое давление, располагаясь преимущественно в его внеклеточных пространствах.
Железо, цинк, марганец, селен входят в состав энзимов хрусталика: железо – в цитохром С, выполняющий детоксирующие функции; цинк – в карбоангидразу; марганец – в декарбоксилазу яблочной кислоты и играет большую роль в окислительно-восстановительных реакциях хрусталика. С селеном связана активность глутатионпероксидазы, составляющей важное звено в механизмах антиоксидантной защиты хрусталика.
При старении хрусталик обедняется минеральными веществами, и его зольный остаток у людей 61–80 лет уменьшается до 3 %. При этом в помутневшем хрусталике снижается концентрация магния, цинка, марганца, а также калия. Напротив, концентрация натрия, хлора, кальция, железа и меди увеличивается. Это сопровождается нарушением процессов активного транспорта глюкозы, аминокислот, катионов, необходимых для нормального функционирования хрусталика.
Органические вещества. В составе органических веществ хрусталика важную группу составляют адениловые нуклеотиды (АТФ, АДМ, АМФ), входящие в состав фосфорсодержащих соединений. Система АДФ – АТФ действует как универсальный переносчик энергии. К другой важной группе органических фосфатов хрусталика относятся динуклеотиды: никотинамидадениндинуклеотид (НАД) и никотинамидадениндинуклеотидфосфат (НАДФ), участвующие в качестве кофакторов в окислительно-восстановительных реакциях.
Помимо вышеописанных нуклеотидов в хрусталике обнаруживают также такие нуклеотиды, как гуанозин – моно-, ди-, трифосфат и другие нуклеотиды с иными азотистыми основаниями, участвующие в ферментном синтезе РНК и ДНК, полисахаридов и фосфолипидов.
Из нефосфорилированных органических веществ для метаболизма хрусталика имеют большое значение:
• Трипептид глутатион, в составе которого имеется цистеин, содержащий свободную сульфгидрильную группу. Глутатион выступает в качестве поставщика кислорода. Его рассматривают также и как природный антиоксидант, защищающий хрусталик от повреждающего воздействия. Исключительно важная роль глутатиона заключается и в сохранении прозрачности хрусталика. Его синтез осуществляется непосредственно в хрусталике из составляющих его аминокислот с использованием энергии АТФ.
• Аскорбиновая кислота полностью поступает в хрусталик из камерной влаги, где секретируется цилиарным телом. Аскорбиновая кислота играет определенную роль в энергопродуцирующих процессах, транспортируя в хрусталик водород. Кроме того, аскорбиновая кислота входит в состав антиокислительной системы хрусталика.
• Витамины: никотиновая кислота, B1, В2, А влияют на митотическую активность эпителия хрусталика; витамин Е рассматривается как возможный антиокислительный фактор, предотвращающий развитие помутнений хрусталика.
При старении хрусталика и развитии его помутнений уменьшается содержание динуклеотидов, глутатиона, аскорбиновой кислоты. Это негативно влияет на окислительно-восстановительные процессы в хрусталике и может способствовать развитию старческой, травматической и осложненной катаракт.
Углеводы. Энергетические потребности хрусталика покрываются в основном за счет «сгорания» углеводов. Основным источником энергии в хрусталике является глюкоза, которая поступает к нему преимущественно из камерной влаги и в меньшей степени из стекловидного тела.
Для снабжения энергией анаболических реакций в хрусталике расщепление глюкозы осуществляется путем аэробного и анаэробного гликолиза, прямого окисления глюкозы (пентозофосфатный путь) и цикла трикарбоновых кислот Кребса. Возможен также сорбитный путь усвоения глюкозы хрусталиком.
Из других углеводов в хрусталике обнаружены также глюкозамин, полисахарид гликоген, сиаловые кислоты, играющие опосредованно определенную роль в дыхании хрусталика.
С возрастом энергетический обмен хрусталика претерпевает определенные изменения. Снижается интенсивность процессов гликолиза и активность его ферментов. Блокирование гликолиза в эксперименте ведет к помутнению хрусталика. Помимо гликолиза, с возрастом уменьшается также интенсивность процессов прямого окисления глюкозы и расщепления глюкозы в цикле трикарбоновых кислот Кребса. При старческой катаракте происходит снижение активности ферментов гликолиза, уменьшается продуцирование АТФ в хрусталике вследствие нарушений гликолиза, процессов прямого окисления глюкозы и путем цикла Кребса, что является одной из причин нарушения обмена белков в хрусталике и его помутнений.
Превалирование сорбитного пути усвоения глюкозы хрусталиком лежит в основе развития диабетической катаракты. Накопление сорбита ведет к набуханию хрусталика из-за осмотических нарушений и изменений конформации его белков.
Липиды. Хотя хрусталик и небогат липидами, в нем обнаружены представители всех их основных групп. В состав нейтральных липидов хрусталика и ходят холестерин, гликолипиды – ганглиозиды, фосфолипиды и фосфоглицериды. Почти 40 % липидов – это полярные фосфолипиды, которые входят в состав клеточных мембран хрусталика. Две трети нейтральных и полярных липидов связаны с белками (липопротеиды), а одна треть находится в свободном состоянии.
Жирные кислоты липидов хрусталика являются насыщенными и мононенасыщенными.
Образование липидов в хрусталике идет крайне медленно, преимущественно из глюкозы. При этом наиболее интенсивно синтез липидов осуществляем си в эпителии хрусталика.
С возрастом в хрусталике человека увеличивается в 1,5–2 раза содержание холестерина, жирных кислот, фосфолипидов. Накопление липидов в хрусталике особенно характерно для старческих катаракт.
Другой особенностью развития старческой катаракты являются изменения в отношении холестерин/фосфолипиды, возрастающие до 4,2–4,5 при норме 3,4– 3,5, а также свободные липиды/липопротеиды в сторону нарастания первых, что отражает потерю липидов из организованных мембран клеток и переход их в аморфные, межклеточные отложения.
В итоге следует заметить, что существует три универсальных механизма повреждения мембран и процессе старческого катарактогенеза: это активация перекисного окисления липидов (ПОЛ) с накоплением в хрусталике продуктов свободнорадикального окисления липидов, которые инициируются активными формами кислорода и продуктами фотоокисления триптофана – производными кинуренина.
Второй н третий механизмы старческого катарактогенеза связаны с увеличением активности фосфолипаз и накоплением в мембранах продуктов деградации фосфолипидов, разрушающих мембраны.
Белки составляют 35% массы хрусталика, а их физико-химическое состояние во многом определяют его прозрачность. Белки неоднородны по составу и представлены водорастворимыми α-, β-, γ- кристаллинами (в соотношении 37 : 62 : 1) и нерастворимым альбуминоидом, который у человека рассматривается как производный от α-кристаллина.
Молекула кристаллинов построена из нескольких субъединиц, каждая из которых содержит SH-группу. В образовании альбуминоидов из кристаллинов играет роль возникновение дополнительных водородных связей между составляющими его субъединицами.
Помимо перечисленных белков, в хрусталике в незначительном количестве присутствуют сложные белки – гликопротеиды, располагающиеся в мембранах волокон, между волокнами, а также в капсуле хрусталика; коллагеноподобные белки капсулы, соединенные с углеводным компонентом; рибо- и дезоксирибонуклеотиды, липопротеиды, фосфопротеиды и хромопротеиды. В состав различных белков хрусталика входят все обычно встречающиеся аминокислоты (около 20) без количественного их преобладания.
Характерной чертой белков хрусталика является их высокая органоспецифичность при низкой видовой специфичности. В хрусталике, как и в любой ткани, на протяжении всей жизни происходит постоянное обновление белков, их синтез и расщепление. Местом наиболее интенсивного синтеза белков в хрусталике является эпителий и периферические слои коры. Необходимые для синтеза белка аминокислоты поступают в хрусталик из камерной влаги. Синтез белковых молекул осуществляется обычным путем. Вначале с помощью матричной РНК из аминокислот образуются полипептидные цепи, затем с помощью как ковалентных, так и нековалентных взаимодействий между полипептидами формируются белковые молекулы.
Обмен белков хрусталика включает и расщепление их до пептидов при помощи протеиназ, а затем далее до аминокислот под действием пептидаз. В нормальном хрусталике синтез белков количественно преобладает над их расщеплением. Образующиеся при протеолизе аминокислоты пополняют аминокислотный фонд хрусталика.
Преобладающей среди свободных аминокислот в хрусталике является глютаминовая кислота, играющая ключевую роль в окислительном расщеплении аминокислот. В хрусталике установлены процессы переаминирования, окислительного дезаминирования аминокислот и функционирует цикл образования мочевины, а содержание свободного аммиака в нем даже выше, чем в других тканях.
В возрастном онтогенезе в хрусталике происходит снижение доли растворимых и нарастание доли нерастворимых белков. По мере старения уменьшается биосинтез кристаллинов, β-кристаллины заменяются α-кристаллинами, и последние превращаются в альбуминоид во всех слоях хрусталика, особенно и его ядре. В белках хрусталика снижается содержание SH-группы, появляются пигментированные соединения.
Убыль растворимых белков происходит не только путем их перехода в нерастворимые, но и за счет просачивания низкомолекулярных форм через капсулу за пределы хрусталика.
При старческих катарактах качественные изменения белков хрусталика сходны с имеющимися изменениями при старении организма, но выражены более резко. Кроме того, при старческих катарактах возможно и образование растворимых агрегатов белков с очень высокой молекулярной массой, которые также снижают прозрачность хрусталика. Образованию таких агрегатов белков способствуют и процессы окисления сульфгидрильных групп растворимых протеинов с образованием дисульфидных сшивок между полипептидами.
Нуклеиновые кислоты. Содержание нуклеиновых кислот в хрусталике очень низкое, особенно это касается ДНК, что объясняется в первую очередь небольшим количеством ядросодержащих клеток.
РНК хрусталика представлена тремя фракциями: РНК-микросом и митохондрий; растворимая фракция; фракция, связанная с альбуминоидом. Образование альбуминоидной РНК происходит из растворимой фракции. И спою очередь, в растворимой фракции присутствуют также специфические транспортные РНК для аминокислот: лейцина, глицина, валина, гистидина, глютамина, лизина.
Информационная РНК в хрусталике представлена двумя типами – короткоживущая, которая несет информацию для синтеза белка в эпителиальных чистках, и долгоживущая (стабильная), обеспечивающая информацией тот же процесс в волокнах.
Общее содержание РНК в хрусталике уменьшается в направлении от его поверхностных к глубоким слоям. Синтез РНК происходит преимущественно в эпителии и молодых волокнах экваториальной зоны хрусталика. При этом растворимая РНК синтезируется в 15 раз активнее микросомальной и в 27 раз активней альбуминоидной.
ДНК содержится в ядрах эпителиальных клеток хрусталика и его волокон. Большее количество ДНК содержится в эпителии центральной зоны хрусталика и меньшее – в приэкваториальной. Синтез ДНК может осуществляться во всех зонах эпителия хрусталика.
С возрастом и при старческих катарактах снижается уровень РНК в хрусталике, в основном за счет уменьшения уровня информационной РНК, и в то же время увеличивается количество альбуминоидной РНК.
При старении хрусталика выраженным изменениям в его эпителии подвергается локализация клеток, синтезирующих ДНК.
Ошибки репарации ДНК способствуют помутнению хрусталика. Установлен факт повреждения ДНК хрусталика ионизирующей радиацией, и этот неблагоприятный эффект опосредуется супероксидным анион-радикалом.
В общем виде процессы, ответственные за нативность хрусталика, представляются в следующем виде.
1. Биофизические и биохимические компоненты, повреждение которых ведет к нарушению его оптических свойств:
• нарушение целостности и взаиморасположения хрусталиковых волокон;
• изменения структуры и проницаемости мембран;
• денатурация, полимеризация, протеолиз, фотолиз и пострибосомальная модификация белков;
• перекисное окисление, расщепление липидов;
• полимеризация, деполимеризация углеводов, гликолизирование белков.
2. Процессы, поддерживающие нативность молекулярных и надмолекулярных структур хрусталика:
• транспорт ионов и метаболитов – аминокислот, витаминов, коферментов, липидов, углеводов;
• биосинтетические процессы, в результате которых образуются белки, липиды, углеводы, глутатион и др.;
• биоэнергетические процессы – окислительно-восстановительные реакции аккумуляции энергии в виде АТФ и других высокоэргических соединений;
• активность энзиматических систем защиты белков, липидов и регенерации восстановленной формы глутатиона – катапаза, супероксидисмутаза, глутатионпероксидаза, глутатион-γ-трансфераза, глутатионредуктаза;
• группа ферментов детоксикации – система цитохрома-450, урокиназа;
• ферменты, восстанавливающие НАДФ.
Основные формы возрастной катаракты (кортикальная, ядерная и субкапсулярная) имеют свои биохимические особенности, которые могут быть перечислены в следующем виде:
а) в механизме образования кортикальных помутнений лежат нарушения равномерного распределения белков в хрусталиковых волокнах, конфигурации белков, плотности их упаковки в волокнах; образование за счет дисульфидных связей высокомолекулярных белковых агрегатов; повышение проницаемости мембран хрусталика и осмотические нарушения, приводящие к усилению обводненности хрусталика; нарушение механизмов транспорта ионов и их энергетического обеспечения;
б) для ядерной катаракты характерны повреждения белков (увеличение размеров молекул, образование нерастворимых высокомолекулярных агрегатов), дезинтеграция мембран волокон, часто сочетающихся с окрашиванием ядpа в желто-коричневый цвет различной степени интенсивности благодаря образованию хромофоров;
в) в генезе формирования субкапсулярных катаракт преобладают осмотические механизмы, нарушения действия «катионового насоса», гидратация хрусталика, набухание и деградация белков, нарушения проницаемости капсулы хрусталика и ее структуры, затрудняющие проникновение метаболитов.
Хрусталик человека является двояковыпуклым и расположен за радужной оболочкой, прикрепляясь к ресничному телу. В своем ложе хрусталик удерживается эластической цинновой связкой и гиалоидохрусталиковой связкой Вигера. В отдельных высказываниях Корнелия Цельса (50-25 г.до н.э.) и Галена (131-201 г.до н.э.) встречаются данные не только о хрусталике, но и о возможных причинах его помутнения. Иоганн Кеплер (1571-1630) предположил о возможной преломляющей роли хрусталика, а Риссо в 1705 году доказал, препарируя глаза умерших, что причиной слепоты может быть помутение хрусталика.
В качестве диоптрического аппарата глаза он воспроизводит на поверхности сетчатки уменьшенную и перевернутую картину рассматриваемого предмета. В то же время хрусталик является для сетчатки световым светофильтром, предохраняющим ее от вредных для нее коротковолновых световых лучей. Абсорбируя в существенной мере синие и фиолетовые лучи, хрусталик помогает уменьшать в глазу хроматичнеские аберрации, превращающие края изображения в цветные.
Помутнения хрусталика, или катаракта, возникают вследствие ряда причин. Разработанные методы хирургического лечения не всегда приводят к восстановлению зрения. Поэтому одним из актуальных вопросов офтальмологии является разработка неинвазивных методов лечения катаракты, для чего необходимы исчерпывающие данные о морфологических особенностях хрусталика и его взаимодействии с окружающими структурами. Это послужило основанием для обоснования цели нашего исследования.
Нами изучены глаза человека в возрасте от 30 до 60 лет с помоцью морфологических методов исследования.
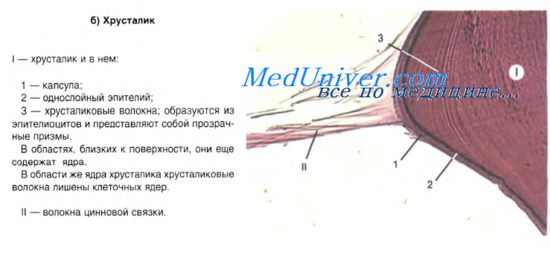
Установлено, что хрусталик состоит: 1) из собственного вещества хрусталика, образованного длинными шестигранными волокнами с двумя широкими и четырьмя узкими поверхностями; 2) из окружающей его эластической капсулы или сумки хрусталика; 3) из эпителия хрусталика, расположенного субкапсулярно на передней поверхности органа и состоящего из одного слоя кубических или плоских клеток. Эпителий покрывает лишь внутреннюю поверхность передней капсулы, поэтому носит название эпителия передней сумки. Клетки его имеют шестиугольную форму. У экватора клетки приобретают вытянутую форму и превращаются в хрусталиковые волокна. Образование волокон совершается в течение всей жизни, что приводит к увеличению хрусталика. Однако чрезмерного увеличения хрусталика не происходит, так как центральные, более старые волокна, теряют воду, уплотняются и постепенно в центре образуют компактное ядро. Плазматическая мембрана клеток содержит поры, облегчающие прохождение через них питательных веществ. Ядро окружено двухконтурной мембраной с порами. Наружный ее слой является продолжением эндоплазматического ретикулума. В цитоплазме присутствуют многочисленные рибосомы, митохондрии небольшого размера и обычного строения, элементы комплекса Гольджи, плотные лизосомы. Видны пиноцитозные вакуоли, центриоли, микротрубочки. Помимо актина в эпителиальных клетках хрусталика человека обнаружены тубулин и виметин.
Функция эпителия хрусталика заключается в волокнообразовании. Дифференцировка клеток морфологически выражается в прогрессирующем удлинении клеток, основания которых смещаются к экватору на заднюю капсулу, а вершины растут кпереди от экватора по направлению к переднему полюсу. Поэтому волокнообразующий эпителий переходит непосредственно в более молодые волокна хрусталика, причем синтез в хрусталиковых волокнах осуществляется преимущественно на основе диплоидной организации их ядер.
Центральная, более плотная часть хрусталика - его ядро - состоит из меридионально расположенных волокон с зазубренными краями и лишенных ядер. Волокна, составляющие более мягкую периферическую субстанцию снабжены ядрами, имеют гладкие контуры и расположены несколько спирально. Вещество, связывающее волокна, накапливается на передней и задней сторонах хрусталика в виде трехлучевой хрусталиковой звезды. Здесь происходит стык хрусталиковых волокон. При этом волокна, берущие начало от центра звезды, заканчиваются на противоположной стороне в конце луча другой звезды, и, наоборот. Таким образом, волокна не охватывают всей половины хрусталика. Хрусталиковые звезды располагаются так, что лучи одной проходят в промежутке между лучами другой. У человека хрусталиковые звезды неправильно-многолучевые.
Капсула хрусталика состоит из близких к коллагену склеропротеинов и полисахаридов, но содержит в виде следов также глутатион и нуклеотиды. Ей свойственно двоякое преломление. В электронном микроскопе обнаруживается фибриллярная структура капсулы хрусталика.
Хрусталик прикреплен к ресничному телу при помощи цилиарной связки, которая состоит из гомогенных и нерастяжимых волокон, начинающихся от базальной мембраны цилиарного эпителия и прикрепляющихся к хрусталиковой капсуле по обе стороны экватора. Экваториальная поверхность хрусталика вместе с передними и задними волокнами ресничного пояска ограничивает пространство, на меридиональных срезах имеющее треугольную форму. Это пространство носит название канала Петита или Ганновера. Фактически канала здесь нет, так как ресничный поясок образован не сплошными пластинками, а отдельными нитями.
Имеется мнение, что ресничный поясок не только подвешивает хрусталик, но и обеспечивает поступление к нему питательных веществ от отростков цилиарного тела. Для офтальмохирурга при экстракапсулярной экстракции катаракты существенный интерес представляет асимметрия крепления ресничного пояска. Поскольку с медиальной стороны зона его крепления уже, чем с латеральной, наиболее опасной при хирургическом вмешательстве является экваториальная зона шириной 2,2 мм с латеральной стороны и 0,9 мм с медиальной от экватора.
Передняя поверхность хрусталика контактирует со зрачковым краем радужной оболочки и в области зрачка омывается влагой передней камеры глаза. На остальном протяжении передняя поверхность хрусталика, его экватор и небольшой приэкваториальный участок омываются внутриглазной жидкостью задней камеры. Большая же часть задней поверхности хрусталика соприкасается со стекловидным телом, отделяясь от него узкой капиллярной щелью - захрусталиковым пространством Бергера. По наружному краю захрусталиковое пространство ограничивается гиалоидокапсулярной связкой фиксирующей хрусталик к стекловидному телу. Эта связка Вигера состоит из тончайших фибрилл, выходящих из пограничной мембраны стекловидного тела. При натяжении задней порции волокон ресничного пояска при операции тракция может передаваться на переднюю гиалоидную мембрану стекловидного тела и сетчатку, вызывая их травматизацию.
Кровеносные сосуды и нервы в хрусталике отсутствуют, поэтому он лишен чувствительности, а трофическое обеспечение осуществляется путем осмоса.
Работа представлена на научную международную конференцию «Современные проблемы экспериментальной и клинической медицины», Бангкок, Паттайа (Тайланд), 20-30 декабря 2008 г. Поступила в редакцию 10.12.2008.
Промежуточные филаменты хрусталика
• Хрусталик глаза содержит два специфических белка промежуточных филаментов, СР49 и филензин, которые входят в VI группу гомологичных последовательностей
• Эти специфические белки промежуточных филаментов сохранили свою структуру в процессе эволюции позвоночных
Клеткам хрусталика глаза позвоночных свойственны крайне жесткие критерии программы дифференцировки, что необходимо для последующего правильного функционирования ткани. Клетки должны:
• Обладать достаточной упругостью и в то же время эластичностью, которая допускает аккомодацию хрусталика.
• Сохранять полную прозрачность для того, чтобы не затемнять хрусталик.
• Сохранять перечисленные качества на протяжении всей жизни организма.
В клетках хрусталика содержатся два специфических белка, которые образуют «филаменты типа бус». Такое название они получили за характерный профиль поверхности, видимый в электронном микроскопе (остальные промежуточные филаменты имеют гладкую поверхность).
Эти белки называются СР49 (или факинин) и филензин. Они относятся к белкам VI типа. Оба белка имеют структурные характеристики, свойственные другим типам белков, однако их последовательности не гомологичны ни одной последовательности среди белков остальных групп.
Одним из наиболее существенных различий является наличие у них «знаковой» последовательности, свойственной белкам промежуточных филаментов, а именно последовательности на конце спирального участка. У CP49 на конце стержнеобразного домена находится последовательность Tyr-His-Gly-Ile-Leu-Asp-Gly-Glu, в то время как у других белков промежуточных филаментов на этом месте присутствует консервативная последовательность Tyr-Arg-Lys-Leu-Leu-Glu-Gly-Glu.
Гены, кодирующие СР49 и филензин, сильно отличаются от генов, кодирующих другие белки промежуточных филаментов. Такое своеобразие этих генов дает им селективное преимущество перед соответствующими генами других клеток, поскольку высоко гомологичные белки хрусталика сохраняются у всех позвоночных.
Последовательность, соответствующая CP49/факинину, обнаружена в геноме сростночелюстной рыбы Fugu ribripes, позвоночного организма, эволюционно очень далеко отстоящего от млекопитающих. Консервативность структуры проявляется даже у специфической последовательности, находящейся на конце спирального участка СР49/факинина.
Белки клеток хрусталика проявляют особые свойства в силу ряда причин. Во-первых, они обеспечивают развитие максимально прозрачного хрусталика. Во-вторых, поскольку хрусталик должен функционировать в течение всей жизни организма, его полимерные структуры должны обладать исключительной биохимической устойчивостью с тем чтобы не допустить деградации белков и изменений их структуры, которые могут привести к нарушению его физических или оптических характеристик и к утрате функции.
Считается, что специфические последовательности и морфологические особенности CP49 и филензина каким-то образом способствуют поддержанию свойств этих белков. В экспериментах на животных с измененным характером экспрессии белков филаментов хрусталика показано развитие последовательных нарушений функционирования хрусталика глаза. Также показано, что доминантные мутации в CP49 являются причиной развития ранней семейной катаракты у человека.
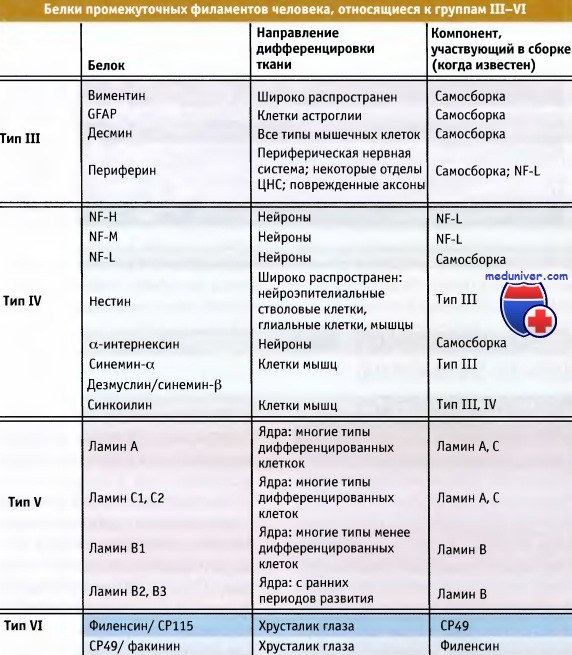
Белки промежуточных филаментов человека, объединенные в типы III—IV по принципу гомологии.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Анатомия и эмбриология хрусталика в норме
а) Анатомия хрусталика глаза. Хрусталик, так же как и роговица, обладает двумя основными оптическими свойствами: прозрачностью и преломляющей силой, определяемыми его структурой. Он представляет собою прозрачное двояковыпуклое бессосудистое образование, состоящее из уникальным образом дифференцированных эпителиальных клеток. Он расположен сразу же за радужкой и удерживается позади зрачка зонулярными волокнами цилиарного тела.
При рождении экваториальный диаметр хрусталика составляет 6,5 мм, а максимальный передне-задний размер между полюсами — 3,5 мм. Хрусталик упакован в коллагеновую капсулу, являющуюся базальной мембраной лежащего под ней однослойного кубического эпителия. Кубические клетки экваториальной зоны хрусталика делятся в течение всей жизни и формируют вторичные веретенообразные волокна хрусталика.
Эти новые волокна экваториальной области постепенно изменяют форму хрусталика от почти сферической у плода до эллипсоидной двояковыпуклой у детей и в молодом возрасте. При рождении имеются эмбриональное и фетальное ядра. Фетальное ядро отграничено от эмбрионального Y-образным швом спереди и перевернутым Y-образным швом сзади. В ходе дальнейшего развития формируются более поверхностные зоны ядра. Образующиеся после рождения волокна хрусталика формируют взрослое ядро.
Таким образом, ядро хрусталика образовано плотно упакованными хрусталиковыми волокнами (утратившими ядра эпителиальными клетками), плотность которых в периферической коре хрусталика ниже. Дальнейший рост хрусталика сопровождается изменениями в основном передне-заднего размера; в молодом возрасте хрусталик имеет стабильный экваториальный диаметр приблизительно 9 мм и передне-задний размер 5 мм.
б) Эмбриология хрусталика глаза. Хрусталик развивается из утолщения поверхностной эктодермы, покрывающей глазной пузырь. Эта «хрусталиковая плакода» у эмбриона человека появляется на 26-27 день развития. У эмбриона курицы глазной пузырь посредством экстрацеллюлярного матрикса плотно фиксируется к поверхностной эктодерме. Фиксация митотически активной поверхностной эктодермы вызывает повышение плотности клеток, удлинение и утолщение плакоды.
Адгезия глазного пузыря к хрусталиковой плакоде постепенно приводит к выравниванию сетчатки и хрусталика по зрительной оси, однако прямой клеточный контакт между базальной мембраной глазного пузыря и поверхностной эктодермой отсутстует. Затем хрусталиковая плакода инвагинирует и формирует хрусталиковую ямку, а затем и хрусталиковый пузырек.
Отслойка хрусталикового пузырька — первый этап формирования переднего сегмента глаза (33 день). Она сопровождается миграцией эпителиальных клеток через роговично-хрусталиковый стебель, некрозом клеток и разрушением базальной мембраны. Нарушение этих процессов при воздействии экзогенных/эндогенных факторов приводит к развитию аномалий переднего сегмента. Отделившийся хрусталиковый пузырек выстлан одним слоем цилиндрических эпителиальных клеток, окруженных базальной пластинкой, будущей капсулой хрусталика.
Образование первичных хрусталиковых волокон происходит в эпителиальных клетках, выстилающих заднюю поверхность хрусталикового пузырька. Этот процесс стимулируется прилегающим зачатком сетчатки. Таким образом, цитодифференциация хрусталика определяется зачатком сетчатки. Первичные волокна хрусталика заполняют просвет хрусталикового пузырька. В результате удлинения клеток хрусталика, прилегающих к сетчатке, формируется эмбриональное ядро хрусталика. Клетки передней части хрусталика, лежащие вблизи зачатка роговицы, сохраняют кубическую форму и однослойную организацию и формируют эпителий хрусталика, который сохраняет митотическую активность в течение всей жизни и впоследствии становится источником хрусталиковых волокон. В области экватора хрусталика эпителиальные клетки дифференцируются во вторичные хрусталиковые волокна (хрусталиковый водоворот).
Эти волокна удлиняются спереди и сзади и внедряются над первичными волокнами хрусталика. В области экватора их толщина наибольшая. Это вызывает увеличение преимущественно экваториального диаметра хрусталика плода. Вторичные хрусталиковые волокна сходятся спереди и сзади и формируют Y-образные швы.
Зонулярные волокна развиваются из беспигментного цилиарного эпителия в течение пятого месяца гестации. Основным компонентом цилиарной связки является гликопротеин фибриллин. Существует две его изоформы, фи-бриллин-1 и фибриллин-2. Полимеры фибриллина-1, без участия значительного количества эластина, образуют структурный каркас растяжимых микрофибрилл. Они образуют параллельные пучки и формируют зонулярные волокна.
При нарушении структуры или функции этих богатых фибриллином микрофибрилл наблюдается зонулярная дисфункция, приводящая к эктопии хрусталика.5 Мутации гена фибриллина-1 (FBN1) вызывают синдром Марфана (MFS — Marfan’s syndrome), вероятно, некоторые формы синдрома Вейля-Маркезани (WMS — Weill-Marchesani syndrome) и различные другие фенотипы фибриллинопатий 1 типа, при которых часто встречается эктопия хрусталика. Наоборот, мутации гена фибриллина-2 (FBN2) могут вызывать синдром Билса, фенотипически сходный с синдромом Марфана, но редко сопровождающийся эктопией хрусталика (см. ниже).
Сосудистая оболочка хрусталика (tunica vasculosa lentis — TVL) — сосудистая сеть из системы гиалоидной артерии, лежащая на задней поверхности хрусталика и параллельные радиальные сосудистые анастомозы с кольцевыми кровеносными сосудами в латеральных отделах. Она опутывает развивающийся хрусталик и питает его в период, когда продукция водянистой влаги и формирование переднего сегмента еще не начались. Эта внутриглазная сосудистая сеть начинает развиваться на первом месяце гестации, достигает максимального развития на втором-третьем месяце и начинает регрессировать к четвертому месяцу. К рождению она почти полностью исчезает.
Персистирующая сосудистая оболочка хрусталика часто выявляется у недоношенных новорожденных, не регрессировавшие сосудистая оболочка хрусталика и гиалоидные артерии могут вызывать различные аномалии, обозначаемые групповым названием «персистирующая фетальная сосудистая сеть» (persistent fetal vasculature — PFV).
Раннее развитие переднего сегмента.
(А) Примерно пять недель развития эмбриона человека, хрусталиковый пузырек отделился от поверхностной эктодермы,
клетки нервного гребня мигрируют вокруг глазного бокала и между поверхностной эктодермой и развивающимся хрусталиком.
(Б) В течение седьмой недели из слоя мезенхимы развиваются выстланная эндотелием строма роговицы и передняя строма радужки.
Этот процесс дифференцировки идет одновременно с разделением двух слоев и формированием передней камеры между развивающимися роговицей и радужкой.
(В) Листок мезенхимы, перекидывающийся через будущий зрачок, сохраняется до седьмого месяца гестации.
Из краев глазного бокала образуется задний эпителий радужки и эпителий цилиарного тела. (А) Формирование глазного пузыря на латеральной стенке диэнцефалона. Зрительный стебель соединяет глазной пузырь с передним мозгом. (9,5 дней гестации мыши, соответствуют 26 дням гестации человека).
(Б) Инвагинация глазного пузыря и образование хрусталикового пузырька (начало 10,5 дней гестации мыши, соответствует 28 дням гестации человека).
(В) Инвагинация хрусталиковой ямки, формирование из инвагинированного глазного пузыря двухслойного глазного бокала (конец 10,5 дней гестации мыши, соответствует 32 дням гестации человека).
(Г) Закрытие эмбриональной хориоидальной щели, формирование хрусталикового пузырька и первичного стекловидного тела (12,5 дней гестации мыши, соответствует 44 дням гестации человека).
(Д) Образование слоя нервных волокон, миграция клеток нервного гребня и образование ядерного пояса хрусталика (14,5 дней гестации мыши, соответствует 56-60 дням гестации человека).
(Е) Глаз в конце стадии органогенеза. Хорошо видны роговица, начинающие формироваться радужка, зачатки экстраокулярных мышц и слезная железа.
Стрелками показана зрачковая мембрана (16,5 дней гестации мыши соответствуют >60 дней гестации человека).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Читайте также: