
Какие элементы обеспечивают тактильную чувствительность кожи
Обновлено: 26.04.2024
Тактильная сенсорная система, наряду с проприорецептивной, зрительной и вестибулярной сенсорными системами, «поставляет» головному мозгу информацию о положении и движении тела в пространстве, о положении его отдельных частей. Кроме того, она играет важную роль в ориентации человека в окружающей среде (особенно сильно кожное осязание развивается у слепых и слепоглухонемых, позволяя тем самым таким людям избегать воздействия повреждающего агента).
Благодаря тактильной сенсорной системе осуществляется контакт грудного ребенка с матерью, выполнение различных игровых, образовательных и трудовых операций, интимные отношения между мужчиной и женщиной, ощущение комфорта от носимой одежды. В принципе, такие примеры, доказывающие значение тактильной рецепции, можно перечислять еще и еще. Скажем только одно – нарушение этого вида чувствительности существенно снижает адаптационные возможности человека и лишает его многих радостей жизни. Особое место тактильная сенсорная система занимает у млекопитающих, выполняя жизненно важные функции – тактильное исследование окружающей среды, питание, звукообразование и т.д.
Тактильная чувствительность резко повышена на участках тела, которые покрыты волосами. Это объясняется тем, что волосы играют роль рычажков и усиливают передачу раздражения в несколько раз, а так как 95% поверхности человеческого тела покрыто волосами, на некоторых участках едва заметными, то любое прикосновение к поверхности тела резко усиливается.
Рецепторы осязательного анализатора
Различают следующие основные механорецепторы – свободные неинкапсулированные нервные окончания, свободные нервные окончания волосяных фолликулов, диски Меркеля, тельца Руффини, тельца Мейснера и тельца Пачини. Строение указанных рецепторов различно, они распределены неравномерно и локализуются на разной глубине кожи. Первые два типа рецепторов относятся к первичным (являются окончаниями дендрита чувствительного нейрона), остальные – ко вторичным (представляют собой инкапсулированные специализированные клетки, трансформирующие механическое воздействие в рецепторный потенциал, который передается на дендрит чувствительного нейрона). Рассмотрим отдельные виды рецепторов (рис. 4.1).
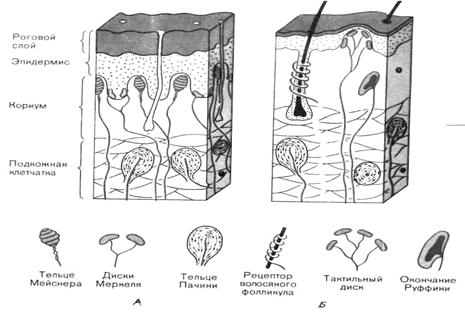
Рис. 4.1. Схема строения и положения механорецепторов в коже, на не покрытых волосами (А) и волосистых (Б) участках кожи.
Свободные неинкапсулированные нервные окончания являются самыми распространенными рецепторами кожи. Они находятся в основном в сосочковом слое дермы – обычно идут вдоль мелких сосудов и представляют собой разветвления дендрита афферентных нейронов. Первоначально их считали рецепторами боли, но в настоящее время их рассматривают как полимодальные рецепторы, отвечающие и на болевые, и на температурные, и на механические стимулы. Это медленно адаптирующиеся рецепторы: продолжают отвечать весь период времени, пока действует стимул.
Свободные нервные окончания волосяных фолликулов также представляют собой разветвления дендрита афферентного нейрона, которые оплетают волосяную сумку. Обычно фолликул получает волокна от нескольких чувствительных нейронов, но в то же время один и тот же дендрит чувствительного нейрона иннервирует несколько волосяных фолликулов. Волос, как было уже отмечено выше, служит рычагом, усиливающим раздражение нервных окончаний, что и объясняет высокую чувствительность волос к прикосновению. Указанные рецепторы реагируют преимущественно на легкое прикосновение и осуществляют пространственное и временное тактильное различение.
Диски (клеточные комплексы) Меркеля представляют собой видоизмененные эпителиальные клетки, с которыми образуют синапсы дендриты афферентных нейронов. Находятся в базальном слое эпидермиса и частично в сосочковом слое дермы в виде небольших скоплений. Особенно много дисков Меркеля в участках кожи с высокой чувствительностью, т.е. в голой коже пальцев рук и на губах. В покрытой волосами коже они лежат в особых колоколообразных тельцах, выдающихся над окружающей кожной поверхностью (в каждом таком образовании, называемом тактильным тельцем Пинкуса-Игго, находится 30-50 клеток Меркеля, связанных с афферентными нервными волокнами). При высоте около 0,1 мм и диаметре 0,2-0,4 мм эти тельца едва различимы невооруженным глазом.
Поскольку диски Меркеля медленно адаптирующиеся рецепторы, то их рассматривают в качестве пропорционального датчика, т.е. генерация рецепторного потенциала в них тем активнее, чем сильнее действует раздражитель. Эти рецепторы рассматривают как рецепторы давления, или силы, поскольку они воспринимают изменение силы механического воздействия.
Тельца (окончания) Руффини располагаются в волосистой части кожи – в глубоких слоях эпидермиса и в сосочковом слое дермы. Представляют собой веретеновидной формы капсулу, образованную плотно переплетенными коллагеновыми волокнами. Внутри такой капсулы содержится жидкость, в которую погружены разветвления дендрита чувствительного нейрона. Как и диски Меркеля, являются рецепторами давления, или силы.
Тельца Мейснера (тактильное мейснеровое тельце) представляют собой капсулу конусовидной или овальной формы. Капсула ориентирована перпендикулярно поверхности кожи. Ее стенки образованы многочисленными пластинчатыми клетками, между которыми параллельно поверхности кожи располагаются терминали дендрита чувствительного нейрона. Эти рецепторы находятся в сосочках собственно кориума, в участках, лишенных волос (кожа ладоней, подошв, пальцев рук и ног, а также губы, сосочки молочных желез и половые органы). В покрытой волосами коже их меньше и здесь они принимают форму рецепторов волосяных фолликулов. Реагируют на скорость изменения силы, т.е. являются датчиком скорости.
Тельца Пачини (пластинчатые тельца, тельца Фатера-Пачини) – наиболее крупные и наиболее распространенные в организме рецепторы, а потому они являются наиболее изученными рецепторами кожи. Располагаются в гиподерме и частично в глубоких слоях дермы. Кроме того, они обнаружены и в сухожилиях и фасциях мышц, в надкостнице, в суставных сумках. Тельце Пачини напоминает луковицу, состоит из наружной капсулы, внутренней колбы и заключенного в нее дендрита афферентного нейрона. Пространство между наружной капсулой и внутренней колбой, а также внутри колбы заполнено жидкостью (ликвором). Возбуждение этих рецепторов происходит при кратковременном (в течение 0,1 мс) смещении капсулы на 0,5 мкм.
Тельца Пачини считаются датчиком ускорения. Благодаря своим свойствам они воспринимают самую незначительную деформацию кожи при соприкосновении с различными предметами и почвой, т.е. с какой скоростью смещается кожа.
Таким образом, большинство тактильных рецепторов локализовано, как правило, в глубоких слоях эпидермиса и сосочковом слое кориума.
Всех их можно разделить на три главных типа:
- рецепторы давления (датчики силы), которые ведут себя как пропорциональные датчики, т.е. генерация рецепторного потенциала в них тем активнее, чем сильнее действует раздражитель. Поэтому их еще называют пропорциональными рецепторами. Это свободные неинкапсулированные нервные окончания, диски Меркля, тельца Руффини;
- рецепторы прикосновения (датчики скорости) реагируют на скорость изменения силы, т.е. скорость вдавления стимула, поэтому осуществляют пространственное и временное тактильное различение. Это тельца Мейсмера и рецепторы волосяных фолликулов;
- рецепторы вибрации (датчики ускорения) – тельца Пачини - реагируют на изменение скорости механического воздействия.
Однако при этом необходимо подчеркнуть, что механические стимулы, обычно действующие на кожу, одновременно возбуждают в той или иной степени несколько типов механорецепторов в зависимости от характера стимула. Поэтому возникающие ощущения нельзя приписать рецепторам одного типа. Соответственно в повседневной жизни трудно определить различия между ощущениями давления и прикосновения.
Принцип работы механорецепторов . Независимо от того, что одни рецепторы реагируют на изменение силы, вторые – на скорость изменения этой силы, а третьи – на вторую производную изменения силы, во всех случаях принцип работы рецептора состоит в том, что под влиянием механического стимула в мембране рецептора изменяется ионная проницаемость, что приводит к появлению рецепторного потенциала. Он вызывает выделение медиатора, что сопровождается появлением генераторного потенциала в дендритах чувствительного нейрона, благодаря которому изменяется частота генерации потенциалов действия.
Различия в восприятии механических стимулов определяются скоростью адаптации тактильных рецепторов. Так, быстро адаптирующиеся рецепторы являются датчиками скорости, или ускорения, а медленно адаптирующиеся – датчиками изменения силы. В то же время скорость адаптации определяется структурой рецепторов – наличие сложно организованной капсулы рецептора повышает скорость его адаптации (укорачивает рецепторный потенциал), поскольку такая капсула хорошо проводит быстрые и гасит медленные изменения давления. Адаптация механорецепторов кожи имеет большое значение – благодаря этому свойству рецепторов мы перестаем ощущать постоянное давление одежды, привыкаем носить на роговице глаз контактные линзы, а на носу – очки и т.д.
Проведение осязательной информации в центральный отдел
Сенсорная информация от механорецепторов кожи передается в кору больших полушарий по нервным волокнам тонкого и клиновидного пучков, образующих лемнисковую систему. Тонкий пучок, или пучок Голля, несет информацию от рецепторов нижних конечностей и нижней части туловища, а клиновидный пучок, или Бурдаха, - от рецепторов верхних конечностей и верхней части туловища. Оба пути состоят из афферентных нейронов (они находятся в спинномозговых узлах), аксоны которых вступают в задние рога спинного мозга и, не прерываясь, направляются в составе задних столбов до тонкого (ядро Голля) и клиновидного (ядро Бурдаха) ядер продолговатого мозга.
Аксоны нейронов этих ядер полностью перекрещиваются на уровне олив, образуя медиальный лемниск (петлю), и, поднимаясь дальше, заканчиваются в специфических ядрах таламуса, которые из-за своего анатомического положения называют вентробазальным комплексом. Нейроны специфических ядер таламуса посылают свои аксоны в соматосенсорные зоны коры больших полушарий (SI и SII). Поражение задних столбов клинически проявляется в потере мышечно-суставного чувства, вибрационной и тактильной чувствительности на стороне поражения при сохранении болевой и температурной чувствительности.
Лемнисковая система проводит точную (по силе и месту воздействия) и сложную (о давлении, прикосновении, вибрации и движении в суставах) информацию с большой скоростью (до 80 км/с).
Для всех составляющих лемнисковой системы (задних столбов спинного мозга, тонкого и клиновидного ядер, таламических ядер и корковых областей) выявлена четкая топографическая организация проекции кожи, т.е. каждый участок кожи контрлатеральной части тела занимает определенную зону, площадь которой зависит от функционального значения этой части тела.
Часть импульсов от рецепторов давления кожи и слизистой передается в кору больших полушарий и по вентральному спиноталамическому тракту, который проходит в составе передних столбов спинного мозга. Однако в спиноталамической системе на уровне таламических ядер правильная соматотопическая организация представительства поверхности тела отсутствует.
Особое место в передаче сенсорной информации от механорецепторов отводится V паре черепно-мозговых нервов - тройничному нерву, который в своих трех ветвях содержит афференты, идущие от лица и полости рта. Он иннервирует кожу, зубы, слизистую полости рта, язык и роговицу. Ко времени рождения тройничная система уже хорошо развита и проводит сенсорные сигналы, которые запускают пищевое поведение. Именно она обеспечивает первое сенсорное знакомство новорожденного с окружающим миром.
Корковый уровень анализа тактильной информации
Информация от нейронов специфических ядер таламуса первоначально поступает в две проекционные соматосенсорные зоны коры больших полушарий (SI и SII). В частности, информация от нейронов вентробазального комплекса направляется (контрлатерально) в первую проекционную зону, которая у приматов и человека находится в постцентральной извилине (SI). От нейронов задней группы ядер таламуса информация преимущественно поступает (контрлатерально и ипсилатерально) во вторую проекционную соматосенсорную зону коры (SII), которая расположена в области сильвиевой (латеральной) борозды (рядом со слуховой зоной). От этих двух проекционных соматосенсорных зон информация поступает в передние и задние ассоциативные зоны коры.
Первая проекционная соматосенсорная область, локализованная в постцентральной извилине (первичные зоны — 1-е и 3-е поля по Бродману, вторичные зоны — 2-е и 5-е поля), является, по сути, местом окончания лемнискового пути и ядром тактильного анализатора. Она отличается от других областей коры очень высокой степенью топографической организации (проекции различных областей поверхности тела на соматосенсорную кору осуществляются по принципу "точка в точку"). Такое явление называют соматотопией, или топографическим представительством. О наличии соматотопии свидетельствует тот факт, что при раздражении поверхности кожи короткими точечными прикосновениями первичный ответ в соматосенсорной коре локализуется в строго ограниченном участке.
Показано, что размеры представительства соответствующих участков поверхности кожи у человека и приматов в постцентральной извилине связаны не с величиной поверхности их тела, а зависят от биологической значимости информации, воспринимающейся тем или иным участком кожи. Например, у человека представительство губ, лица и кистей рук в этой зоне по площади намного больше, чем представительство туловища и нижних конечностей. Это дает специфический рисунок сенсорного гомункулюса — отражения тела человека в постцентральной извилине (рис. 4.2). Характер этого рисунка свидетельствует о высокой чувствительности и тонком различении для этих частей тела, а также их биологической значимости. Так, при осмотре какого-либо предмета, когда нам надо выяснить форму, наличие шероховатостей и т.д., мы поглаживаем этот предмет, т.е. касаемся его поверхности кожей нашей ладони.
Когда кора в области SI разрушена повреждением или удалена с терапевтической целью, возникает дефицит восприятия. Стимуляция кожи еще может быть воспринята как таковая, но способность локализовать ее и распознать пространственные детали раздражителя нарушается. Так, например, больные не могли на ощупь отличить овал от параллелепипеда (в клинике это нарушение носит название астерогнозис). Степень дефицита зависит от размеров поврежденного участка коры. Здесь снова выступает соматотопическая организация. Однако надо сказать, что после достаточно долгого промежутка времени подобные нарушения становятся слабее. Такое улучшение, по-видимому, обусловлено способностью других областей коры (например, 5-го поля теменной коры, соседнего с SI) принять на себя функции SI.
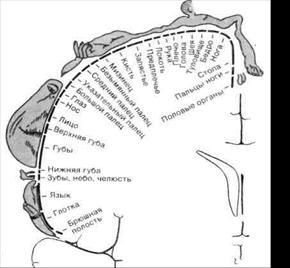
Рис.4.2. Схема чувствительного гомункулюса (соматотопическая организация соматосенсорной коры SI человека).
Разрез полушарий (на уровне постцентральной извилины) во фронтальной плоскости. Обозначения показывают пространственное представительство поверхности тела в коре, установленное на основании локальной стимуляции мозга бодрствующих больных.
Анализ информации от тактильных рецепторов в первой соматосенсорной зоне осуществляется нейронами, объединенными в вертикальные колонки, которые можно рассматривать как своеобразные функциональные единицы, или блоки коры. Каждая такая колонка, получая информацию от рецепторов одной и той же модальности, находящихся на одном и том же рецептивном поле кожи, проводит этот анализ с участием специализированных нейронов, число которых в колонке достигает 10 5 . Каждый из этих нейронов «настроен» на определенный признак, наличие которого в поступающей информации вызывает возбуждение соответствующего нейрона. Благодаря деятельности колонок мозг получает информацию обо всех свойствах стимула, воздействующего на соответствующий участок кожи.
Во вторую соматосенсорную зону коры больших полушарий (SII), расположенную в области сильвиевой борозды вблизи от слуховой зоны (40-е и 51-е поля), поступают импульсы от тактильных рецепторов кожи «своей» и противоположной стороны. Эта зона содержит точное и детальное представительство поверхности тела, как и первая соматосенсорная зона, с тем различием, что проекции обеих половин тела во второй соматосенсорной зоне полностью перекрываются, благодаря чему происходит объединение и сравнение информации поступающей от правой и левой половины тела, т.е. имеет место билатеральное соматотопическое представительство. Как полагают, SII специально играет роль в сенсорной и моторной координации активности на двух сторонах тела (например, хватание или ощупывание обеим руками). Не исключено, что вторая соматосенсорная зона может, кроме того, осуществлять контроль над афферентной передачей сигналов в таламических ядрах.
От первичных и вторичных проекционных зон коры информация от тактильных рецепторов поступает в передние (фронтальные) и задние ассоциативные зоны коры, благодаря которым завершается процесс восприятия, т.е. происходит опознание образа (акцепция сигнала). Это реализуется с участием специальных нейронов («бабушкиных» нейронов), проходящих «обучение» в процессе индивидуального развития человека.
В целом роль соматосенсорной зоны коры состоит в интегральной оценке соматосенсорных сигналов, во включении их в сферу сознания, полисенсорный синтез и в сенсорное обеспечение выработки новых двигательных навыков. Удаление или повреждение соматосенсорных зон коры приводит к нарушению способности локализовать тактильные ощущения, а их электростимуляция вызывает ощущение давления, прикосновения, вибрации и зуда.
Соматовисцеральная система перерабатывает сенсорную информацию нескольких модальностей, получая ее от кожи, мышц, суставов, внутренних органов и кровеносных сосудов. Она содержит разные типы рецепторов: механорецепторы, терморецепторы, хеморецепторы, осморецепторы, но-цицепторы. Поступающая информация передается по афферентным волокнам в составе чувствительных и смешанных нервов, перерабатывается раздельными проводящими путями и переключательными ядрами, а затем доставляется в соматосенсорную кору, занимающую постцентральные извилины мозга и организованную топически.
Тактильная чувствительность
Часть соматовисцеральной системы, обеспечивающая чувство осязания, включает несколько разновидностей механорецепторов кожи, представленных свободными нервными окончаниями либо инкапсулированными, т. е. заключенными в капсулу из соединительной ткани или видоизмененных клеток эпидермиса (рис. 17.4). Свободные нервные окончания иннервируют волосяные фолликулы пушковых волос, покрывающих большую часть тела человека, а также грубых волос, растущих на голове, в подмышечных впадинах, на лобке, а у мужчин еще и на лице. Свободные нервные окончания волосяных фолликулов являются механоре-цепторами и возбуждаются при смещении волос или их подергивании. Другая разновидность свободных нервных окончаний имеется в эпидермисе и в сосочковом слое дермы, большинство из них являются ноцицепторами или терморецепторами, но некоторые принадлежат к механорецепторам, которые специфически реагируют на слабое околопороговое раздражение. Предполагается, что при раздражении этой разновидности рецепторов возникают ощущения щекотки и зуда.

Рис. 17.4. Схема распределения механорецепторов в коже человека. Свободные нервные окончания имеют высокий порог раздражения и слабо реагируют на изменение интенсивности стимула. Быстро адаптирующиеся рецепторы (тельца Пачини, рецепторы волосяных фолликулов) служат датчиками скорости действующих стимулов, а медленно адаптирующиеся рецепторы (диски Меркеля, тельца Руффини) являются датчиками интенсивности действующего раздражителя. Наличие нескольких разновидностей рецепторов позволяет передавать афферентные сигналы о разных свойствах одного и того же раздражителя.
Среди инкапсулированных окончаний различают тельца Пачини, Мейсснера, Руффини, диски Меркеля, тактильные тельца Пинкуса—Игго, колбы Краузе. В зависимости от строения и формы капсулы нервные окончания подвержены наиболее сильному воздействию либо в результате давления действующим перпендикулярно раздражителем, либо вследствие бокового смещения капсулы, которая играет роль механического преобразователя энергии внешних стимулов. Большинство инкапсулированных рецепторов содержится в лишенной волос коже пальцев рук и ног, ладоней и подошв, лица, губ, языка, сосков и половых органов, где они распределены с различной плотностью и на разной глубине. Тельца Пачини имеются также в сухожилиях, связках и брыжейке.
Механорецепторы кожи различаются по скорости адаптации к действующему раздражителю. Быстроадаптирующиеся (фазные) рецепторы возбуждаются только в момент смещения кожи и волос и служат датчиками скорости воздействия стимула. Это свойство присуще тельцам Мейснера, рецепторам волосяных фолликулов и особенно тельцам Пачини, способным реагировать на изменения скорости продолжающего свое действие стимула. Медленно адаптирующиеся (тонические) рецепторы не прекращают генерировать потенциалы действия при продолжительном действии раздражителя, если он оказывает давление на кожу: такие рецепторы служат датчиками интенсивности действующего стимула (тельца Руффини, диски Меркеля).
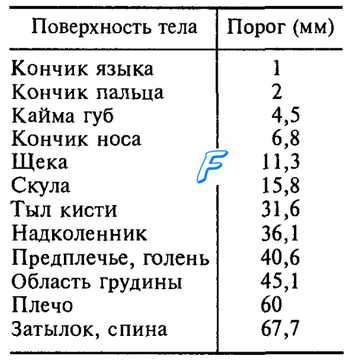
Таблица 17.1. Пространственный двухточечный порог в разных участках тела
Площадь рецептивных полей сенсорных нейронов, иннервирующих тельца Мейснера и диски Меркеля, составляет в среднем около 12 мм2, а у нейронов с окончаниями в виде телец Пачини и Руффини она на порядок больше. Рецептивные поля различающихся своими рецепторами сенсорных нейронов перекрываются, поэтому при действии на кожу комплекса стимулов одновременно возбуждаются разные виды рецепторов, что позволяет ощущать все динамические и статические свойства такого комплекса. Обработка и анализ информации сигналов от различных рецепторов происходит на высших уровнях сенсорной системы, формирующих комплексное восприятие действующих на поверхность тела стимулов. Плотность меха-норецепторов в разных участках кожи не одинакова, чем определяются разные показатели пространственного дифференциального порога, т. е. наименьшего расстояния между двумя точками, раздражение каждой из которых ощущается раздельно (табл. 17.1). Приведенные в таблице данные не следует считать эталоном, поскольку дифференциальная чувствительность различается у разных людей.
Инкапсулированные рецепторы иннервируются миелинизированными волокнами первичных сенсорных нейронов, которые проводят нервные импульсы в ЦНС со скоростью около 30—70 м/с. Немиелинизированные волокна передают потенциалы действия от свободных нервных окончаний со значительно меньшей скоростью — около 1 м/с, поэтому ощущение действующего на них стимула возникает относительно позже. Центральные отростки первичных сенсорных нейронов входят в спинной мозг в составе задних корешков и разделяются в задних рогах спинного мозга на коллатерали. Восходящие коллатерали достигают переключательных ядер заднего столба продолговатого мозга, откуда специфическая информация передается на противоположную сторону мозга по лемнисковому пути, поступает к проекционным ядрам таламуса, а затем в соматосенсорную кору, с участием которой формируется чувство осязания.
Кожная чувствительность. Методы оценки кожной чувствительности
Во внутриутробный период и непосредственно после рождения ребенка чувствительность кожи имеет важное значение для анализа изменений, происходящих во внешней среде. Восприятие контактных раздражений осуществляется главным образом кожей, которая чувствительна к болевым, тактильным и температурным воздействиям.
Болевая чувствительность на механические раздражения у новорожденных развита слабо, особенно у недоношенных, но в течение первых недель жизни она заметно возрастает. У новорожденных порог раздражения для боли значительно выше, чем у детей более старшего возраста. Доношенные новорожденные уже в 1—2-й день после рождения отвечают на укол иглой средней силы отчетливой болевой реакцией в виде локального движения, крика или сморщивания лица. Область лица у новорожденных и грудных детей особенно чувствительна к болевым раздражениям. У недоношенных новорожденных даже сильные уколы иглой могут не вызвать реакции неудовольствия. Постепенно, с возрастом, чувствительность к болевым раздражениям возрастает, и ответная реакция носит генерализованный характер. После 2 мес жизни общая реакция на болевое раздражение уменьшается, а локальные реакции становятся более отчетливыми. Ребенок стремится избавиться от раздражителя, отодвигая раздражаемый участок, совершает движение конечностями или сильным плачем сигнализирует окружающим. К концу 1-го года жизни он может локализовать место раздражения, болевая чувствительность в этот период уже хорошо развита.
Болевая чувствительность к электрическому току у детей раннего возраста ниже, чем у взрослых и детей старшего возраста. Пониженная чувствительность к раздражению электрическим током сохраняется почти до 6-летнего возраста.
Функциональное развитие тактильной чувствительности опережает развитие остальных органов чувств. К концу 2-го месяца внутриутробной жизни устанавливаются первые реакции на тактильное раздражение. В течение последующих месяцев они завершают свое развитие и у плода 7 мес сходны с реакциями у новорожденного ребенка.

Тактильная чувствительность у детей раннего возраста выражена довольно хорошо, но, как и у взрослого, разные части тела неодинаково чувствительны к прикосновению. Наиболее чувствительны лицо, кожа рук, стоп и менее — кожа предплечья, лопаток, груди, живота, спины, бедер и голеней.
В первые 2 мес у детей отсутствует реакция на щекотание: она появляется обычно после 2 мес и с 9 мес имеется у всех детей. Вначале она вызывается с кожи подмышечной впадины, затем — со стоп и шеи в виде неудовольствия, плача, защитных движений, а позднее в форме положительной эмоциональной реакции, улыбки, смеха.
Температурная чувствительность у новорожденного хорошо развита. Он рзагирует как на понижение, так и на повышение температуры. Наиболее выражена ответная реакция при разнице между температурой раздражителя и температурой гела 6—7°. При этом новорожденные сильнее реагируют на снижение температуры, чем на ее повышение.
Болевую чувствительность у новорожденных и грудных детей проверяют, нанося раздражение иглой или электрическим током на определенные участки тела. Ответная реакция в виде отдергивания конечностей или общего беспокойства ребенка свидетельствует о сохранности болевой чувствительности.
Тактильную чувствительность исследуют, нанося на кожу раздражение мягким предметом (ватой, кисточкой). В ответ на него возникают общая и местная двигательные реакции. Так, при поглаживании кончика носа появляются сосательные движения, прикосновение к средней части спинки носа приводит к прищуриванию глаз, поднятию крыльев носа и углов рта; раздражение корня языка и нёба вызывает рвотные движения; раздражение круговой мышцы рта вызывает вытягивание губ (хоботковый рефлекс), а прикосновение к векам, конъюнктиве, ресницам — смыкание глаз. Поглаживание внутренней стороны согнутого мизинца приводит к его разгибанию. При продолжающемся раздражении разгибаются и остальные пальцы (кроме большого). Раздражение внутренней поверхности ладони вызывает хватательный рефлекс.
Температурную чувствительность у грудных детей можно исследовать, раздражая различные участки тела пробирками с холодной и теплой водой. На эти раздражения дети отвечают общим беспокойством или местными двигательными реакциями. Слабые температурные раздражения вызывают сокращение мышц лица и движения ног. Более сильные приводят к генерализованной двигательной реакции, сморщиванию лица, крику.
Все вышесказанное свидетельствует о том, что в раннем детском возрасте можно получить только общие представления о расстройстве чувствительности. Определить границы и тип чувствительных расстройств практически невозможно. Поэтому исследование чувствительности у детей раннего возраста не имеет такого значения, как у взрослых.
Сенсорные рецепторы. Типы сенсорных рецепторов и раздражители

а) Типы сенсорных рецепторов и раздражители, которые они воспринимают. В таблице выше представлен список и классификация большинства сенсорных рецепторов тела. Согласно этой таблице, выделяют пять основных типов сенсорных рецепторов:
(1) механорецепторы, реагирующие на механическое сжатие или растяжение самого рецептора или смежных с ним тканей;
(2) терморецепторы, воспринимающие изменения температуры: одни из них реагируют на холод, другие — на тепло;
(3) ноцицепторы (болевые рецепторы), реагирующие на повреждение ткани независимо от природы повреждения (физической или химической);
(4) электромагнитные рецепторы, воспринимающие свет на сетчатке глаза;
(5) хеморецепторы, которые обнаруживают вкус во рту, запах в носу, уровень кислорода в артериальной крови, осмоляльность жидкостей тела, концентрацию углекислого газа и, возможно, другие химические факторы нашего тела.
В статьях на сайте мы обсудим функцию нескольких типов рецепторов, прежде всего периферических механорецепторов, для иллюстрации некоторых общих принципов работы рецепторов. Другие рецепторы обсуждаются в главах, посвященных соответствующим сенсорным системам, в составе которых они функционируют. На рисунке ниже показаны некоторые механорецепторы, расположенные в коже или глубоких тканях тела.
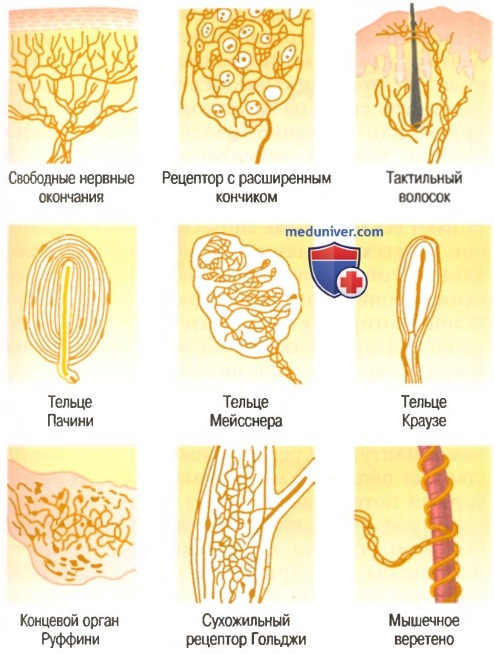
Некоторые типы соматических рецепторов
б) Специфическая чувствительность рецепторов. Прежде всего, необходимо ответить на вопрос, почему разные типы сенсорных рецепторов реагируют на различные типы раздражителей? Ответом на этот вопрос является специфическая чувствительность рецепторов. Это значит, что каждый тип рецептора высокочувствителен к определенному типу стимула, для восприятия которого он предназначен, и практически нечувствителен к другим типам сенсорных стимулов.
Так, палочки и колбочки глаза высокочувствительны к свету, но почти совсем не реагируют на нормальные диапазоны изменений температуры, давления на глазные яблоки или химических показателей крови. Осморецепторы супраоптических ядер гипоталамуса обнаруживают самые незначительные изменения осмоляльности внеклеточных жидкостей тела, но пока неизвестны случаи их реакции на звук.
Наконец, рецепторы боли в коже почти никогда не стимулируются обычным прикосновением к коже или давлением на нее, но реагируют очень активно в тот момент, когда осязательные стимулы становятся достаточно сильными, чтобы повредить ткани.
в) Модальность ощущения. Принцип меченой линии. Отличительную особенность каждого из переживаемых нами ощущений (боль, прикосновение, свет, звук и т.д.) называют модальностью ощущения. Однако, хотя модальности ощущений различны, нервные волокна передают только импульсы. Справедлив вопрос: как возбуждение разных нервных волокон ведет к развитию ощущений различной модальности?
Ответ заключается в том, что каждый чувствительный тракт заканчивается в определенном участке центральной нервной системы, и тип ощущения, испытываемого при стимуляции нервного волокна, определяется тем участком нервной системы, к которому от него придет возбуждение. Например, если раздражается болевое волокно, человек чувствует боль независимо от того, стимул какого типа возбуждает волокно.
Это может быть электрическое раздражение самого волокна, его перегревание, раздавливание или стимуляция болевого рецептора при повреждении тканевых клеток. Во всех этих случаях человек почувствует боль. Аналогично, если тактильное волокно стимулируется электрическим раздражением тактильного рецептора или любым другим способом, человек чувствует прикосновение, поскольку по тактильным волокнам информация достигает определенных чувствительных областей головного мозга. Точно так же волокна от сетчатки глаза заканчиваются в зрительных областях мозга, слуховые пути от уха заканчиваются в слуховых областях, а температурные пути оканчиваются в температурных областях.
Таким образом, специфичность нервных путей способствует развитию ощущений только одной модальности. Эту особенность называют принципом меченой линии.
Видео физиология анализаторов - общая схема - профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Кожные анализаторы тактильной чувствительности определяют тактильные ощущения, которые делятся на ощущения прикосновения, давления, вибрации и зуда.
Причина их возникновения в раздражении рецепторов, представляющих из себя свободные концы нервных сплетений, либо специальные нервные образования: тельца Мейснера на поверхности кожи, лишенной волос, и тельца Пачини в глубоких слоях кожи. Волоски на коже представляют собой рычаги, которые увеличивают силу воздействия чего-либо на кожу.

а - разрез Фатер-Пачиниева тельца кожи человека: 1 - внутренняя колбочка; 2 - нервное волокно. б - разрез тельца Мейснера из сосочка кожи пальца человека: 1 - эпителий; 2,3 - нервные волокна; 4 - капсула.[1]

1 — свободные нервные окончания в эпидермисе; 2 — осязательные тельца Меркеля (главным образом в эпидермисе): 3 — осязательное тельце Мейсснера; 4 — нервное сплетение вокруг волосяной луковицы; 5 — чувствительное к холоду тельце (или колба) Краузе; 6 — фатерпачиниево тельце, чувствительное к давлению.
Тактильные рецепторы располагаются в определённых точках соприкосновения. Месторасположение этих точек определяется раздражением, которое наносится тонким волоском прибора - эстезиометра. Если взаимодействие волоска с кожей минимально, то ощущение прикосновения появится при условии, что кончик волоска совпадёт с такой точкой.
Количество точек прикосновения различается в зависимости от участка кожи. Так максимум их наблюдается в кончиках пальцев и языке. Сами тактильные ощущения соединены с волокнами, по которым идётвозбуждение от тактильных рецепторов. Появляются тактильные ощущения, в связи с возбуждением коры в области задней центральной извилины, которая представляет собой корковый конец кожного анализатора. Отличающиеся участки кожи представлены в коре пространственно различающимися точками, но элементарное соответствие между поверхностью кожи и площадью корковой проекции отсутствует. Так, например, в коре больше всего рецепторов пальцев рук.
Пространственная локализация тактильных ощущений тоже различается: на кончике языка две разные точки воспринимаются на расстоянии 1 — 2 мм. На спине и плечах это расстояние увеличивается до 50 — 60 мм. Если выразить величину среднего порога чувствительности кожи к давлению в абсолютных цифрах, то она колеблется в пределах от 1,13 до 4,3 г на 1 мм 2 . При нагревании кожи чувствительность повышается, при охлаждении соответственно уменьшается.[2]
Локализация тактильных раздражений регистрируется весьма точно. Вопрос этот изучался Вебером. Прикасаясь к коже циркулем и разводя его ножки больше или меньше, он отмечал, на каком расстоянии между ножками регистрируются раздельные ощущения. На кончике языка ощущения раздельно регистрировались при расстоянии в 1,1 мм между ножками циркуля, на поверхности пальцев — 2,2 мм, на средней линии шеи и спины — 67,7 мм. Различать тактильные раздражения можно не только по месту приложения раздражителя, но и по его силе.
Ощущения давления появляются из-за сильного действия раздражителя на кожу, что происходит из-за деформации кожного покрова. Если давление распределяется равномерно, то такое ощущение не появляется. Важное значение принадлежит скорости деформации кожного покрова.
А вот вибрация возникает из-за ритмичных раздражений рецепторов. Высшего развития вибрационная чувствительность достигает у глухих, и в определённой степени восполняет слух.
Температурные ощущения представляют собой отображение уровня тепла тела и появляются при воздействии на кожу предметов с температурой отличной от температуры кожи. Терморецепторы могут подвергаться раздражению, кроме прямого контакта, ещё и по причине лучистого теплообмена между кожей и предметом.
Температурные ощущения разделяются на ощущения тепла и холода.
Тепловые ощущения появляются, если температура выше температуры тела, тогда раздражаются специальные рецепторы тепла, под которыми обычно понимают тельца Руффини. Xолодовые ощущения возникают при температуре ниже температуры тела, что связывается с раздражением специальных рецепторов холода, вероятно колб Краузе.
Специализацию тепловых и холодовых рецепторов можно доказать существованием отдельных тепловых и холодовых точек. Для этого применяют специальный термоэстезиометр, состоящий из трубки и термометра. Тонкое окончание даёт возможность наносить точечные тепловые раздражения. Тепловые и холодовые точки аналогично реагируют и на раздражение током.
Количество тепловых и холодовых точек так же зависит от участка кожи, причем оно подвергается изменению из-за раздражителя, воздействующего на рецептор. Например, при обогреве кожи количество тепловых точек увеличивается. Происходит это благодаря рефлекторной настройке рецептора, находящегося в зоне задней центральной извилины.
Особенности температурных ощущений зависят и от удельной теплоемкости предмета. Железо и дерево, нагретые до одной и той же температуры, вызывают разные эффекты - железо кажется горячее.[3]
Точка отсчёта, от которой зависит возникновение холодовых и тепловых ощущений, может сдвигаться. Так, если погрузить одну руку в сосуд с горячей, а другую руку в сосуд с холодной водой, то при последующем погружении обеих рук в сосуд со средней температурой воды в каждой руке возникнут разные ощущения: рука, находившаяся в сосуде с холодной водой, воспримет воду с средней температурой как теплую, а находившаяся в сосуде с горячей водой—как холодную.
Возникновение температурных ощущений связывается с работой корковой части кожного анализатора, поэтому его можно вызывать условно-рефлекторным путем. Если тепловому раздражению подвергнуть кожу кисти руки следом за действием света, то после ряд сочетаний применение света вызывает ощущение тепла, причем с расширением сосудов руки. Температурные ощущения в ответ на условный раздражитель возникают и при анестезии кожи - когда рецепторы кожи выключены.
Развитие и улучшение морфофизиологических функций и самого организма в целом зависят от их способности к дальнейшему развитию, что имеет по большей часи генетическую основу и крайне важно для достижения необходимых показателей физической и умственной работоспособности. При этом способность к выполнению физической работы может возрастать, но до определенных пределов. В отличие от неё умственная деятельность практически не имеет ограничений в своем формировании. У каждого организма есть свои конкретные резервные возможности. Постоянна мышечная деятельность даёт возможность с помощью совершенствования физиологических функций раскрыть резервы, само существование которых было неизвестно. При этом адаптированный к нагрузкам организм обладает большими резервами, использует их более продуктивно. Человек с относительно высокими морфофункциональными показателями физиологических систем и генов обладает увеличенной способностью к выполнению значительных по мощности, объему, интенсивности и продолжительности физических нагрузок. Особенностями морфофункционального состояния разных систем организма, появившихся в результате двигательной деятельности, стали физиологическими показатели тренированности.[4]
Основной задачей упражнения является сохранение здоровья и работоспособности на нужном уровне за счет активизации восстановительных процессов. В ходе упражнений улучшаются высшая нервная деятельность, функции центральной нервной, нервно-мышечной, сердечно-сосудистой, дыхательной, выделительной и других систем, обмен веществ и энергии, а также системы нейрогуморального регулирования.
Так, к числу показателей тренированности в покое можно отнести:
· изменения в состоянии центральной нервной системы,
· изменения опорно-двигательного аппарата
· изменения функции органов дыхания,состава крови и т.п.
Тренированный организм тратит в покое, меньше энергии, чем нетренированный. Упражнения накладывает особый отпечаток на организм, вызывая в нем как морфологические, так физиологические и биохимические перестройки. Все они направлены на обеспечение высокой активности организма при выполнении работы.
Реакции на стандартные нагрузки у тренированных лиц характеризуются следующими особенностями:
1. показатели работы функциональных систем в начале работы оказываются выше, чем у нетренированных;
2. в процессе работы уровень физиологических сдвигов менее высок;
3. период восстановления короче.
При одной и той же работе тренированные спортсмены расходуют меньше энергии, чем нетренированные. У первых меньше величина кислородного запроса, меньше размер кислородной задолженности, но относительно большая доля кислорода потребляется во время работы. Следовательно, одна и та же работа происходит у тренированных с юношей долей участия аэробных процессов, а у нетренированных — аэробных. Вместе с тем во время одинаковой работы у тренированных ниже, чем у нетренированных, показатели потребления кислорода, вентиляции легких, частоты дыхания.
Тренированный организм выполняет стандартную работу более экономно, чем нетренированный. Тренировка определяет адаптивные изменения в организме, которые вызывают экономизацию всех физиологических функций. Одна и та же работа по мере развития тренированности становится менее утомительной.
Читайте также: