
Перечислить какие ощущения воспринимают кожные рецепторы
Обновлено: 26.04.2024
Иннервация кожи: нервные окончания, клетки Меркеля, тельца Руффини, Мейснера, Пачини
а) Чувствительные единицы. Любое нервное волокно, разветвляясь, дает начало нервным окончаниям одного вида. Стволовое нервное волокно и его нервные окончания, выполняющие одни и те же физиологические функции, представляют собой чувствительную единицу. В совокупности с исходным униполярным нейроном чувствительная единица аналогична двигательной единице, описанной в отдельной статье на сайте.
Область, раздражение которой приводит к возбуждению чувствительной единицы, называют рецепторным полем. Чем больше размер рецепторного поля, тем меньшей остротой сенсорной чувствительности обладает данная область: например, в верхней части руки рецепторные поля занимают площадь 2 см 2 , в области запястья — 1 см 2 , на подушечках пальцев — 5 мм 2 .
Чувствительные единицы переплетаются между собой, за счет чего становится возможным одновременное восприятие одним участком кожи разных видов чувствительности.
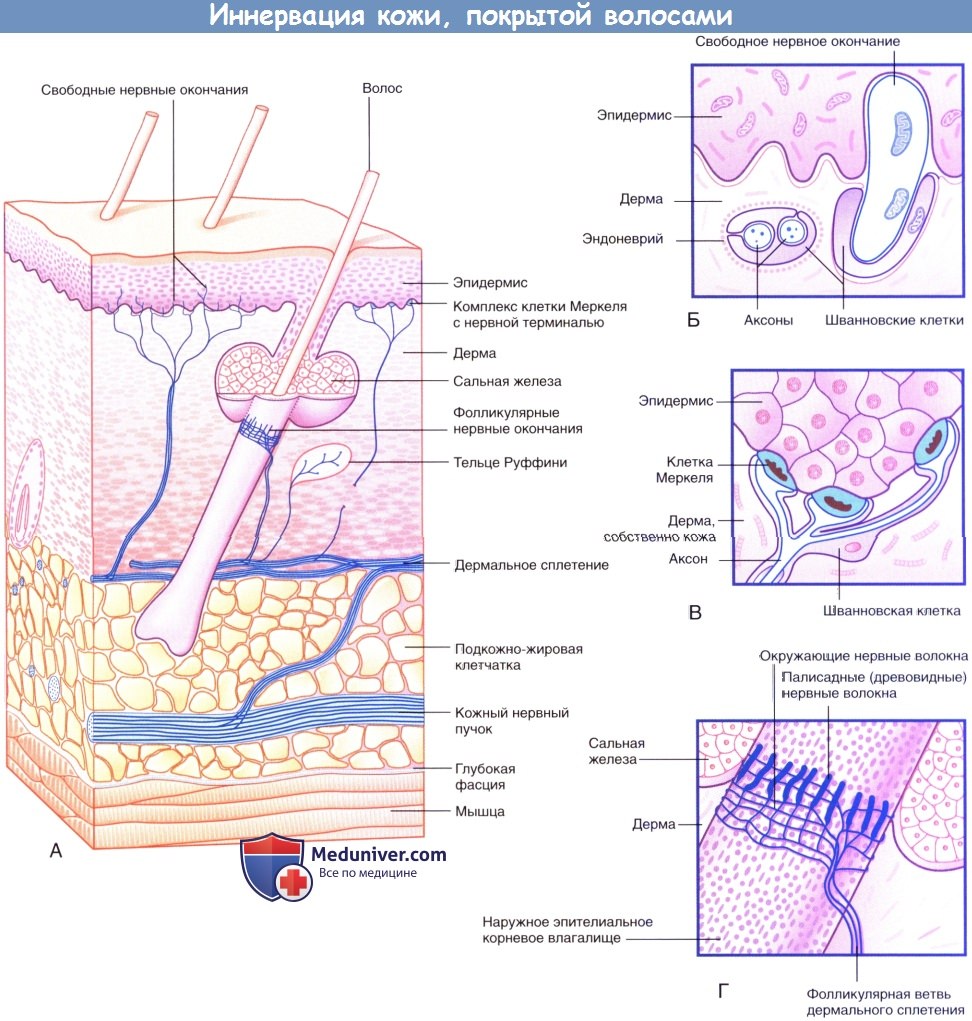
Иннервация кожи, покрытой волосами.
(А) Три морфологических типа чувствительных нервных окончаний в коже, покрытой волосами.
(Б) Свободные нервные окончания в базальном слое эпидермиса.
(В) Комплекс клетки Меркеля с нервной терминалью.
(Г) Палисадные и циркулярные нервные окончания на поверхности наружного корневого влагалища волоса.
б) Нервные окончания:
1. Свободные нервные окончания. По мере приближения к поверхности кожи многие чувствительные нервные волокна утрачивают периневральную, а затем и миелиновую оболочку (в случае ее наличия). Впоследствии нервные волокна разветвляются и формируют субэпидермальное нервное сплетение. Аксон освобождается от оболочек, сформированных шванновскими клетками, что позволяет ему, разветвляясь между коллагеновыми пучками дермы, образовывать дермальные нервные окончания, а внутри эпидермиса — эпидермальные нервные окончания.
Функции. Некоторые чувствительные единицы со свободными нервными окончаниями являются терморецепторами, иннервирующими расположенные на поверхности кожи «тепловые точки» или «холодовые точки». Кроме того, в коже существуют два основных типа ноцицепторов (рецепторов болевой чувствительности), которые также имеют свободные нервные окончания: а-дельта-механоноцицепторы и полимодальные С-ноцицепторы. А-дельта-механоноцицепторы иннервируются тонкими миелинизированными волокнами Аδ-типа и воспринимают существенную механическую деформацию кожи (возникающую, например, при щипке пинцетом). Полимодальные С-ноцицепторы реагируют на болевые стимулы разного вида — механическую деформацию, сильное нагревание или охлаждение (это характерно лишь для некоторых рецепторов), воздействие химических раздражителей. Именно эти рецепторы отвечают за реализацию аксон-рефлекса.
2. Фолликулярные нервные окончания. Нервные окончания волосяного фолликула представлены палисадными нервными волокнами, образованными обнаженными терминалями миелинизированных нервных волокон, расположенными на поверхности наружного корневого влагалища волосяных фолликулов ниже уровня сальных желез, а также циркулярными нервными окончаниями. Каждая фолликулярная единица иннервирует несколько волосяных фолликулов и образует множественные перекресты. Фолликулярные единицы — быстро адаптирующиеся: они возбуждаются при изменении положения волос, однако при сохранении этого положения возбуждения не происходит. Человек, одеваясь, чувствует давление одежды, но затем за счет быстрой адаптации вскоре перестает ощущать ее прикосновение. Иннервация волос у других млекопитающих организована сложнее. Иннервация волосяных фолликулов осуществляется тремя типами механорецепторов, каждый из которых передает информацию определенным структурам мозга, что свидетельствует о важности выполняемой ими чувствительной функции.
3. Комплексы клетки Меркеля с нервной терминалью. Нервная терминаль, расширяясь в области базального слоя эпидермальных гребешков и бороздок, образует комплекс с осязательным тельцем овальной формы — клеткой Меркеля. Комплексы клетки Меркеля с нервной терминалью — медленно адаптирующиеся. В ответ на продолжительное давление (например, при удержании ручки или ношении очков) эти комплексы непрерывно генерируют нервные импульсы. Комплексы клетки Меркеля с нервной терминалью особенно хорошо распознают края удерживаемых в руке предметов.
4. Инкапсулированные нервные окончания. Капсулы описанных ниже свободных нервных окончаний состоят из трех слоев: наружный слой представлен соединительной тканью, средний — периневральным эпителием, а внутренний — видоизмененными шванновскими клетками (телоглией). Инкапсулированные нервные окончания являются механорецепторами, преобразующими механическое воздействие в нервный импульс.
• Тельца Мейснера в большом количестве находятся в подушечках пальцев и расположены вблизи бороздок эпидермиса. Тельца представляют собой клетки овальной формы, внутри которых аксоны располагаются зигзагообразно между уплощенными клетками телоглии. Тельца Мейснера— быстро адаптирующиеся, вместе с медленно адаптирующимися комплексами клетки Меркеля с нервной терминалью они обеспечивают точное восприятие текстур (например, текстуры ткани одежды или поверхности дерева), а также рельефных поверхностей (например, шрифта Брайля). Такие кожные рецепторы способны воспринимать изменение рельефа поверхности даже на высоту 5 нм.
• Тельца Руффини присутствуют как на гладкой коже, лишенной волос, так и на коже с волосами. Они воспринимают плавные скользящие касательные прикосновения и являются медленно адаптирующимися. Внутреннее строение телец сходно со строением сухожильных органов Гольджи: аксоны образуют разветвления в центральной части телец, представленной коллагеновыми волокнами.
• Тельца Пачини по величине соответствуют размерам рисового зерна. В области кисти имеется около 300 телец, которые преимущественно сконцентрированы на боковых участках пальцев и ладони. Тельца Пачини расположены подкожно, близко к надкостнице. Несколько слоев периневрального эпителия внутри соединительнотканной капсулы расположены овально и по форме напоминают луковицу в разрезе. В центральной части тельца Пачини несколько пластинок телоглии окружают единичный аксон, который, попадая в тельце, утрачивает миелиновую оболочку. Тельца Пачини — быстро адаптирующиеся рецепторы преимущественно вибрационной чувствительности. Эти структуры особенно восприимчивы к вибрации костной ткани: большое количество телец расположено в надкостнице длинных трубчатых костей.
Тельца Пачини генерируют один или два нервных импульса при сдавлении и столько же — при прекращении воздействия. В коже ладоней тельца Пачини функционируют по групповому принципу: более 120 телец активируются одновременно, когда человек берет в руку какой-либо предмет (например, апельсин), и когда отпускает его. В связи с этим тельца Пачини считают «детекторами событий» в ходе манипуляций предметами.
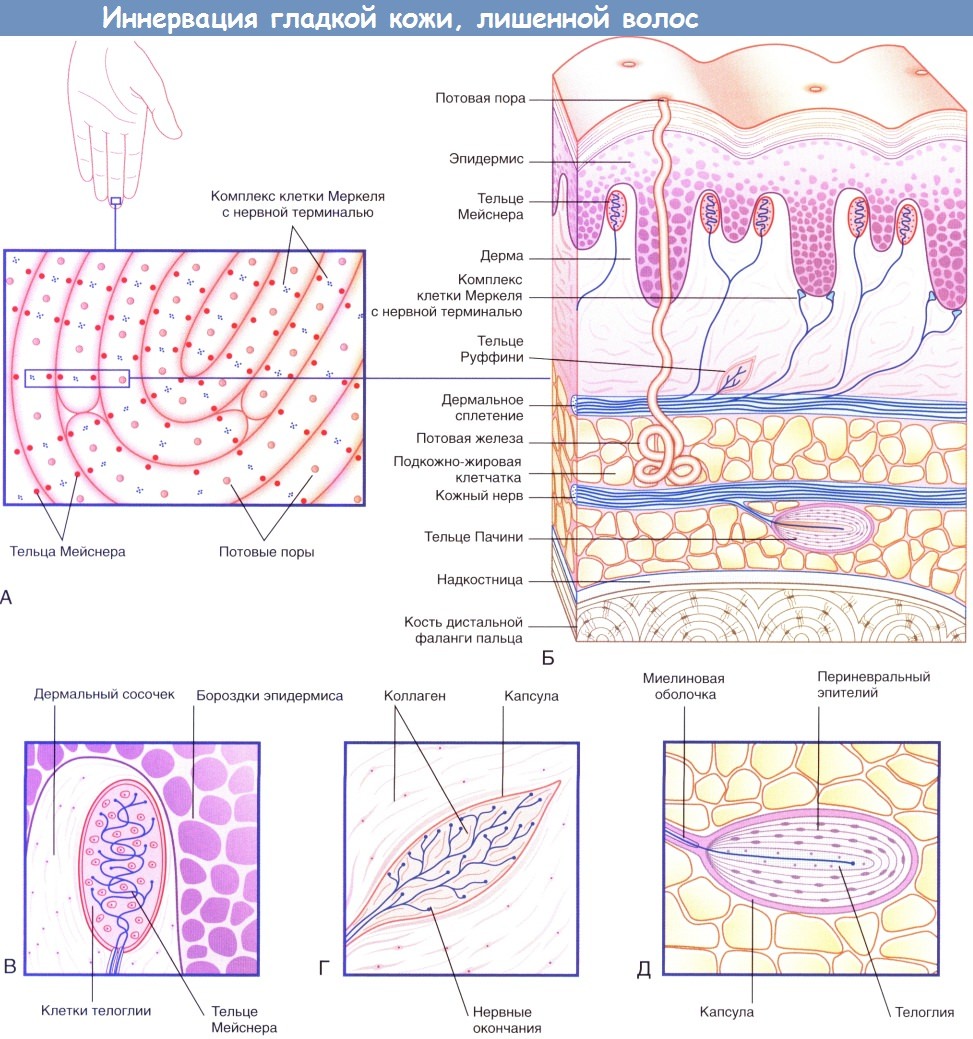
Иннервация гладкой кожи, лишенной волос.
(А) На подушечках пальцев располагаются нервные окончания двух видов.
(Б) На схеме строения участка кожи с изображения (А) представлены четыре типа чувствительных нервных окончаний.
(В) Тельца Мейснера.
(Г) Тельца Руффини.
(Д) Тельца Пачини.
Специалисты по физиологии чувствительности выделяют следующие виды рецепторов, локализованных в коже пальцев.
• Комплексы клетки Меркеля с нервной терминалью — медленно адаптирующиеся рецепторы I типа (MAP I).
• Тельца Мейснера — быстро адаптирующиеся рецепторы I типа (БАР I).
• Тельца Руффини — медленно адаптирующиеся рецепторы II типа (MAP II).
• Тельца Пачини — быстро адаптирующиеся рецепторы II типа (БАР II).
Восприятие ощущений манипуляций с трехмерным предметом вне поля зрения человека в основном обеспечивается за счет мышечных (направляющихся преимущественно от мышечных веретен) и суставных (направляющихся от суставных капсул) афферентных нервных волокон. Кожные, мышечные и суставные афференты независимо друг от друга передают информацию в контралатеральную соматосенсорную зону коры головного мозга. Три различных вида информации объединяются на клеточном уровне в задней части контралатеральной теменной доли, отвечающей за тактильную и визуальную пространственную чувствительность. Тактильную пространственную чувствительность называют стереогнозом. В клинической практике для определения стереогноза пациента просят определить, какой предмет он держит в руках (например, ключ), не смотря на него. Кожные ощущения при периферических нейропатиях описаны в отдельной статье на сайте.
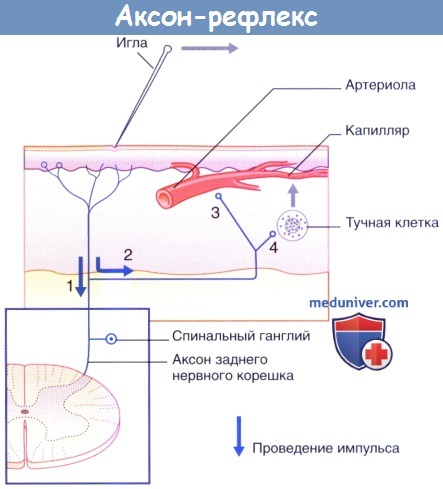
в) Нейрогенное воспаление - аксон-рефлекс. При раздражении чувствительной кожи острым предметом линия контакта практически мгновенно приобретает красный цвет, что обусловлено расширением капилляров в ответ на повреждение кожи. Спустя несколько минут расширение артериол приводит к увеличению зоны гиперемии, а экссудация плазмы из просветов капилляров вызывает формирование бледного отечного валика. Этот феномен представляет собой «тройную реакцию» кожи на раздражение. Формирование зон гиперемии и отечного валика обусловлено аксон-рефлексом чувствительных кожных нервов. Происходящие процессы описаны в соответствии с нумерацией на рисунке ниже.
1. Полимодальные ноцицепторы преобразуют действие болевого раздражителя в нервные импульсы.
2. Аксоны посылают нервные импульсы в центральную нервную систему не только в обычном ортодромном направлении, но и в противоположном антидромном направлении от мест бифуркации к прилежащим участкам кожи. Ответная реакция ноцицептивных нервных окончаний на антидромную стимуляцию проявляется в высвобождении пептидных веществ, среди которых в большом количестве представлена субстанция Р.
3. Субстанция Р связывается с рецепторами на стенках артериол и вызывает их расширение, что приводит к появлению гиперемии.
4. Кроме того, субстанция Р связывается с рецепторами на поверхности тучных клеток, что приводит к высвобождению из них гистамина. Гистамин увеличивает проницаемость капилляров, за счет чего происходит местное накопление тканевой жидкости, обусловливающее возникновение бледного отечного валика.
г) Лепра. Возбудитель лепры — микобактерия, которая проникает в организм человека через мельчайшие повреждения кожи и, распространяясь проксимально по периневрию кожных нервов, вызывает гибель шванновских клеток. Утрата миелиновой оболочки на определенных участках крупных нервных волокон («сегментарная демиелинизация») приводит к нарушению проведения нервных импульсов. Вследствие ответной воспалительной реакции на внедрение возбудителя происходит сдавление всех аксонов, что приводит к валлеровской дегенерации нервов и значительному разрастанию их соединительнотканных оболочек. В результате этого на коже пальцев верхних и нижних конечностей, а также на носу и ушах формируются участки, лишенные чувствительности. Поскольку защитная функция кожной чувствительности нарушается, эти участки становятся более подверженными травматизации, что приводит к повреждению тканей. По мере прогрессирования заболевания возникает двигательный паралич, обусловленный поражением стволов смешанных нервов, расположенных проксимально по отношению к точкам отхождения их кожных ветвей.
д) Резюме. Направляющиеся к коже нервы разветвляются и образуют дермальное нервное сплетение. Чувствительные нервные волокна дермального сплетения разветвляются и перекрывают друг друга. Каждое стволовое нервное волокно и его рецепторы формируют чувствительную единицу. Область, иннервируемую стволовым нервным волокном, называют его рецептивным полем.
К чувствительным единицам со свободными нервными окончаниями относят рецепторы температурной чувствительности, а также механические и температурные рецепторы болевой чувствительности. Рецепторы волосяных фолликулов—быстро адаптирующиеся осязательные механорецепторы, которые активируются только при движении волос. Комплексы клеток Меркеля с нервными терминалями обеспечивают восприятие края предметов, их относят к медленно адаптирующимся.
Инкапсулированные нервные окончания являются механорецепторами. Тельца Мейснера расположены в пространствах между гребешками эпидермиса гладкой кожи, их относят к быстро адаптирующимся. Тельца Руффини—рецепторы растяжения кожи—расположены вблизи ногтей и волосяных фолликулов, их относят к медленно адаптирующимся. Тельца Пачини—подкожные быстро адаптирующиеся нервные окончания, обладающие вибрационной чувствительностью и являющиеся «детекторами событий». На уровне задней части теменной доли коры головного мозга происходит объединение кодированной информации, полученной от кожи, мышц и суставов, что способствует осуществлению тактильного восприятия и стереогностической чувствительности.

78. Где расположены рецепторы, информирующие головной мозг о состоянии опорно-двигательного аппарата?
О состоянии опорно-двигательного аппарата головной мозг информируют импульсы, идущие от рецепторов мышц, сухожилий, связок.
79. Запишите определение.
Осязание – это комплекс ощущений, возникающих при раздражении рецепторов кожи.
80. Перечислите, какие ощущения воспринимают кожные рецепторы.
Прикосновения, давление, боль, холод, тепло.
81. Какие участки тела наиболее чувствительны к тактильным воздействиям?
Поверхность ладоней, кончик носа, подушечки пальцев, кожа над скуловой костью.
82. Рассмотрите рисунок, схематично изображающий строение органа обоняния. Напишите названия структур, обозначенных цифрами.
83. Где в головном мозге расположена обонятельная зона, в которой формируется ощущение запаха?
Обонятельная зона располагается во внутренней поверхности височных долей головного мозга.
84. Перечислите участки, где расположены вкусовые рецепторы.
Вкусовые рецепторы человека расположены в эпителии языка, мягком нёбе и на задней стенке глотки.
85. Подпишите на рисунке названия вкуса (горький, кислый, солёный, сладкий), который лучше всего ощущается в выделенных зонах языка.
86. Какие вещества не имеют вкуса?
Нерастворимые вещества не вызывают раздражения вкусовых рецепторов и поэтому кажутся безвкусными.
87. Дополните схему «Строение периферического отдела вкусового анализатора»
88. Опишите путь, который проходит информация о вкусовых раздражителях от рецепторов до коры больших полушарий.
От рецепторов информация о вкусовых раздражителях в виде нервных импульсов по волокнам языкоглоточного и частично лицевого и блуждающего нервов поступает в средний мозг, ядра таламуса и, затем, на внутреннюю поверхность височных долей коры больших полушарий.
89. Какие ещё рецепторы, помимо вкусовых, участвуют в формировании вкуса?
Тактильные, обонятельные, температурные.
Московский государственный медико-стоматологический университет
Кафедра общей и эстетической стоматологии и кафедра терапевтической стоматологии факультета последипломного образования Московского государственного медико-стоматологического университета
Кафедра общей и эстетической стоматологии и кафедра терапевтической стоматологии факультета последипломного образования Московского государственного медико-стоматологического университета
Вкусовая чувствительность и ее изменения
Журнал: Российская стоматология. 2011;4(2): 15‑19
Крихели Н.И., Гаматаева Д.И., Дмитриева Н.Г. Вкусовая чувствительность и ее изменения. Российская стоматология. 2011;4(2):15‑19.
Krikheli NI, Gamataeva DI, Dmitrieva NG. Gustatory sensitivity and its changes. Russian Stomatology. 2011;4(2):15‑19. (In Russ.).
Московский государственный медико-стоматологический университет
Представлены сведения о строении органа вкуса, описан процесс восприятия вкусового ощущения. Рассмотрены изменения вкусовой чувствительности в норме, при соматических заболеваниях и при беременности. Также приводятся этиологические факторы возникновения нарушения вкусовой чувствительности и метод ее диагностики.
Московский государственный медико-стоматологический университет
Кафедра общей и эстетической стоматологии и кафедра терапевтической стоматологии факультета последипломного образования Московского государственного медико-стоматологического университета
Кафедра общей и эстетической стоматологии и кафедра терапевтической стоматологии факультета последипломного образования Московского государственного медико-стоматологического университета
Орган вкуса и восприятие вкусовой чувствительности
Формирование вкусового ощущения происходит за счет восприятия рецепторами сосочков языка вкусовых раздражителей. За каждый вид раздражителей отвечают определенные зоны языка. Сладкое воспринимают рецепторы кончика языка, кислое - его боковые зоны, горькое - корень языка, соленое - кончик языка и его боковые поверхности [1, 9].

На спинке и боковых поверхностях языка расположены различные группы сосочков (см. рисунок). Рисунок 1. Сосочки языка (схема) по Х. Фениш (цит. по [1]). Нитевидные и конусовидные сосочки воспринимают тактильные и температурные раздражители, а также выполняют механическую функцию; грибовидные, желобовидные (желобоватые) и листовидные - вкусовые. Дополнительно выделяют также чечевицевидные сосочки (papillae lentiformes). Нитевидные сосочки наиболее многочисленные, расположены на дорсальной поверхности языка в поперечном направлении параллельными рядами, плотно прилегают друг к другу, поэтому поверхность языка имеет бархатистый вид. В области корня языка эти ряды нитевидных сосочков повторяют рисунок V-образной бороздки, отделяющей тело языка от его корня. Как установлено рядом исследователей [1, 6], сенсорная функция сосочков обеспечивается нейрофиламентарным протеином и протеином S-100. В состав нитевидных сосочков входят собственная пластинка и эпителий. Каждый сосочек состоит из первичного сосочка, образованного собственной пластинкой, от которого отходят более мелкие вторичные сосочки собственной пластинки. Первичный сосочек покрыт эпителием, который прикрывает и каждый вторичный сосочек. Название «нитевидные сосочки» связано с тем, что участки эпителия, покрывающего вторичные сосочки, по структуре напоминают нити [1]. На верхушке сосочка нити расщеплены в виде «гребешка». Эти участки состоят из вещества, сходного с роговым. Эпителий, покрывающий нитевидные сосочки, ороговевает. Процесс слущивания поверхностного слоя эпителия нитевидных сосочков является выражением физиологической регенерации. При нарушении функции органов пищеварения, при общих воспалительных, инфекционных заболеваниях отторжение поверхностного слоя эпителия нитевидных сосочков замедляется, язык становится «обложенным». То же наблюдается и при гиподинамии языка [1, 6, 9].
Конусовидные сосочки являются особым типом нитевидных сосочков, различающихся анатомическим строением верхушки. В отличие от «гребешка» нитевидных сосочков их нерасщепленная коническая верхушка изогнута кончиком назад.
Грибовидные сосочки расположены на кончике языка, листовидные – по бокам, желобовидные - на корне [1, 3, 9]. Последние окружены валиком, в который открывается секрет мелких белковых желез. Это необходимо, во-первых, для удаления частиц пищи с поверхности сосочка, а во-вторых, при этом растворенное вещество лучше воспринимается рецептором. Именно поэтому при плохой гигиене языка снижается его вкусовая чувствительность.
Грибовидных сосочков меньше, чем нитевидных [1]. Эти сосочки немногочисленны и беспорядочно разбросаны среди нитевидных. Наблюдают их больше на кончике языка, чем в других его участках. Они имеют узкое основание и расширенную вершину. В области эпителия вершины сосочка располагаются вкусовые луковицы. Наибольшее количество их находится в области кончика языка. Каждый грибовидный сосочек образован собственной пластинкой и называется первичным сосочком, от которого в покровный эпителий вдается вторичный сосочек. Однако поверхность их эпителия не повторяет контуров вторичных сосочков в собственной пластинке, как это наблюдается в нитевидных сосочках. Поэтому вторичные сосочки собственной пластинки подводят капилляры близко к поверхности эпителия. Покровный эпителий этих сосочков сравнительно прозрачный, так как он не ороговевает [1]. За счет этого кровь в сосудах, проходящих в высоких вторичных сосочках, просвечивается, придавая им прижизненно красный цвет. В норме грибовидные сосочки бледно-розового цвета и не выделяются выше уровня нитевидных сосочков. В патологии - ярко-красные, гиперплазированные; могут являться инициальными в развитии эрозий и язв на языке. При системных нарушениях за счет повреждений этих сосочков нередко отмечается потеря вкусовой чувствительности [1, 2, 9]. Возможно, они взаимосвязаны с эндокринной системой. В толще самих сосочков определяется большое количество телец Меркеля, ответственных за сенсорную функцию.
В эпителии сосочков выявляются вкусовые луковицы овальной формы, прилежащие своим основанием к базальной мембране и доходящие вершинами до поверхности эпителия, отделяясь от нее 2-3 рядами плоских эпителиальных клеток. В пространстве, разделяющем листовидные сосочки, открываются протоки белковых желез. В области кончика языка имеются смешанные слизистые железы, которые несколькими выводными протоками открываются на нижней поверхности языка [1].
Схема восприятия вкусового ощущения
Вкусовой сосочек - вкусовая почка - мембрана микроворсинок - рецепторные белки - изменяется состав ионов - сигнал о раздражении в кору головного мозга - восприятие вкуса.
Соматовисцеральная система перерабатывает сенсорную информацию нескольких модальностей, получая ее от кожи, мышц, суставов, внутренних органов и кровеносных сосудов. Она содержит разные типы рецепторов: механорецепторы, терморецепторы, хеморецепторы, осморецепторы, но-цицепторы. Поступающая информация передается по афферентным волокнам в составе чувствительных и смешанных нервов, перерабатывается раздельными проводящими путями и переключательными ядрами, а затем доставляется в соматосенсорную кору, занимающую постцентральные извилины мозга и организованную топически.
Тактильная чувствительность
Часть соматовисцеральной системы, обеспечивающая чувство осязания, включает несколько разновидностей механорецепторов кожи, представленных свободными нервными окончаниями либо инкапсулированными, т. е. заключенными в капсулу из соединительной ткани или видоизмененных клеток эпидермиса (рис. 17.4). Свободные нервные окончания иннервируют волосяные фолликулы пушковых волос, покрывающих большую часть тела человека, а также грубых волос, растущих на голове, в подмышечных впадинах, на лобке, а у мужчин еще и на лице. Свободные нервные окончания волосяных фолликулов являются механоре-цепторами и возбуждаются при смещении волос или их подергивании. Другая разновидность свободных нервных окончаний имеется в эпидермисе и в сосочковом слое дермы, большинство из них являются ноцицепторами или терморецепторами, но некоторые принадлежат к механорецепторам, которые специфически реагируют на слабое околопороговое раздражение. Предполагается, что при раздражении этой разновидности рецепторов возникают ощущения щекотки и зуда.

Рис. 17.4. Схема распределения механорецепторов в коже человека. Свободные нервные окончания имеют высокий порог раздражения и слабо реагируют на изменение интенсивности стимула. Быстро адаптирующиеся рецепторы (тельца Пачини, рецепторы волосяных фолликулов) служат датчиками скорости действующих стимулов, а медленно адаптирующиеся рецепторы (диски Меркеля, тельца Руффини) являются датчиками интенсивности действующего раздражителя. Наличие нескольких разновидностей рецепторов позволяет передавать афферентные сигналы о разных свойствах одного и того же раздражителя.
Среди инкапсулированных окончаний различают тельца Пачини, Мейсснера, Руффини, диски Меркеля, тактильные тельца Пинкуса—Игго, колбы Краузе. В зависимости от строения и формы капсулы нервные окончания подвержены наиболее сильному воздействию либо в результате давления действующим перпендикулярно раздражителем, либо вследствие бокового смещения капсулы, которая играет роль механического преобразователя энергии внешних стимулов. Большинство инкапсулированных рецепторов содержится в лишенной волос коже пальцев рук и ног, ладоней и подошв, лица, губ, языка, сосков и половых органов, где они распределены с различной плотностью и на разной глубине. Тельца Пачини имеются также в сухожилиях, связках и брыжейке.
Механорецепторы кожи различаются по скорости адаптации к действующему раздражителю. Быстроадаптирующиеся (фазные) рецепторы возбуждаются только в момент смещения кожи и волос и служат датчиками скорости воздействия стимула. Это свойство присуще тельцам Мейснера, рецепторам волосяных фолликулов и особенно тельцам Пачини, способным реагировать на изменения скорости продолжающего свое действие стимула. Медленно адаптирующиеся (тонические) рецепторы не прекращают генерировать потенциалы действия при продолжительном действии раздражителя, если он оказывает давление на кожу: такие рецепторы служат датчиками интенсивности действующего стимула (тельца Руффини, диски Меркеля).
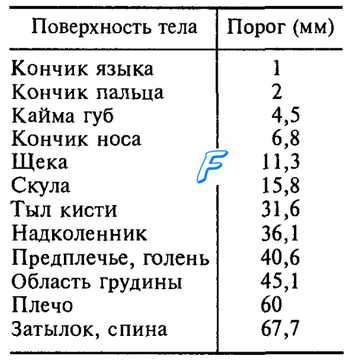
Таблица 17.1. Пространственный двухточечный порог в разных участках тела
Площадь рецептивных полей сенсорных нейронов, иннервирующих тельца Мейснера и диски Меркеля, составляет в среднем около 12 мм2, а у нейронов с окончаниями в виде телец Пачини и Руффини она на порядок больше. Рецептивные поля различающихся своими рецепторами сенсорных нейронов перекрываются, поэтому при действии на кожу комплекса стимулов одновременно возбуждаются разные виды рецепторов, что позволяет ощущать все динамические и статические свойства такого комплекса. Обработка и анализ информации сигналов от различных рецепторов происходит на высших уровнях сенсорной системы, формирующих комплексное восприятие действующих на поверхность тела стимулов. Плотность меха-норецепторов в разных участках кожи не одинакова, чем определяются разные показатели пространственного дифференциального порога, т. е. наименьшего расстояния между двумя точками, раздражение каждой из которых ощущается раздельно (табл. 17.1). Приведенные в таблице данные не следует считать эталоном, поскольку дифференциальная чувствительность различается у разных людей.
Инкапсулированные рецепторы иннервируются миелинизированными волокнами первичных сенсорных нейронов, которые проводят нервные импульсы в ЦНС со скоростью около 30—70 м/с. Немиелинизированные волокна передают потенциалы действия от свободных нервных окончаний со значительно меньшей скоростью — около 1 м/с, поэтому ощущение действующего на них стимула возникает относительно позже. Центральные отростки первичных сенсорных нейронов входят в спинной мозг в составе задних корешков и разделяются в задних рогах спинного мозга на коллатерали. Восходящие коллатерали достигают переключательных ядер заднего столба продолговатого мозга, откуда специфическая информация передается на противоположную сторону мозга по лемнисковому пути, поступает к проекционным ядрам таламуса, а затем в соматосенсорную кору, с участием которой формируется чувство осязания.
Сенсорные рецепторы. Типы сенсорных рецепторов и раздражители

а) Типы сенсорных рецепторов и раздражители, которые они воспринимают. В таблице выше представлен список и классификация большинства сенсорных рецепторов тела. Согласно этой таблице, выделяют пять основных типов сенсорных рецепторов:
(1) механорецепторы, реагирующие на механическое сжатие или растяжение самого рецептора или смежных с ним тканей;
(2) терморецепторы, воспринимающие изменения температуры: одни из них реагируют на холод, другие — на тепло;
(3) ноцицепторы (болевые рецепторы), реагирующие на повреждение ткани независимо от природы повреждения (физической или химической);
(4) электромагнитные рецепторы, воспринимающие свет на сетчатке глаза;
(5) хеморецепторы, которые обнаруживают вкус во рту, запах в носу, уровень кислорода в артериальной крови, осмоляльность жидкостей тела, концентрацию углекислого газа и, возможно, другие химические факторы нашего тела.
В статьях на сайте мы обсудим функцию нескольких типов рецепторов, прежде всего периферических механорецепторов, для иллюстрации некоторых общих принципов работы рецепторов. Другие рецепторы обсуждаются в главах, посвященных соответствующим сенсорным системам, в составе которых они функционируют. На рисунке ниже показаны некоторые механорецепторы, расположенные в коже или глубоких тканях тела.
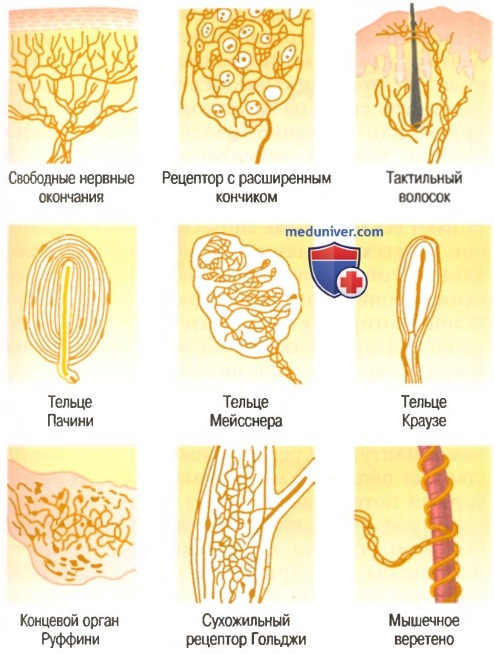
Некоторые типы соматических рецепторов
б) Специфическая чувствительность рецепторов. Прежде всего, необходимо ответить на вопрос, почему разные типы сенсорных рецепторов реагируют на различные типы раздражителей? Ответом на этот вопрос является специфическая чувствительность рецепторов. Это значит, что каждый тип рецептора высокочувствителен к определенному типу стимула, для восприятия которого он предназначен, и практически нечувствителен к другим типам сенсорных стимулов.
Так, палочки и колбочки глаза высокочувствительны к свету, но почти совсем не реагируют на нормальные диапазоны изменений температуры, давления на глазные яблоки или химических показателей крови. Осморецепторы супраоптических ядер гипоталамуса обнаруживают самые незначительные изменения осмоляльности внеклеточных жидкостей тела, но пока неизвестны случаи их реакции на звук.
Наконец, рецепторы боли в коже почти никогда не стимулируются обычным прикосновением к коже или давлением на нее, но реагируют очень активно в тот момент, когда осязательные стимулы становятся достаточно сильными, чтобы повредить ткани.
в) Модальность ощущения. Принцип меченой линии. Отличительную особенность каждого из переживаемых нами ощущений (боль, прикосновение, свет, звук и т.д.) называют модальностью ощущения. Однако, хотя модальности ощущений различны, нервные волокна передают только импульсы. Справедлив вопрос: как возбуждение разных нервных волокон ведет к развитию ощущений различной модальности?
Ответ заключается в том, что каждый чувствительный тракт заканчивается в определенном участке центральной нервной системы, и тип ощущения, испытываемого при стимуляции нервного волокна, определяется тем участком нервной системы, к которому от него придет возбуждение. Например, если раздражается болевое волокно, человек чувствует боль независимо от того, стимул какого типа возбуждает волокно.
Это может быть электрическое раздражение самого волокна, его перегревание, раздавливание или стимуляция болевого рецептора при повреждении тканевых клеток. Во всех этих случаях человек почувствует боль. Аналогично, если тактильное волокно стимулируется электрическим раздражением тактильного рецептора или любым другим способом, человек чувствует прикосновение, поскольку по тактильным волокнам информация достигает определенных чувствительных областей головного мозга. Точно так же волокна от сетчатки глаза заканчиваются в зрительных областях мозга, слуховые пути от уха заканчиваются в слуховых областях, а температурные пути оканчиваются в температурных областях.
Таким образом, специфичность нервных путей способствует развитию ощущений только одной модальности. Эту особенность называют принципом меченой линии.
Видео физиология анализаторов - общая схема - профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Читайте также: