
Как называется слой зернистых эпителиальных клеток
Обновлено: 02.05.2024
Эпителии целомического типа. Эпителии нейроглиального типа.
Целомический тип эпителия мезодермалъного происхождения, как правило, однослойный плоский или призматический, выполняет разграничительную, барьерную, секреторную и другие функции. Целомические эпителии развиваются из материала внутренней выстилки сплахнотома, формирующего целом (вторичную полость тела). Наиболее характерным эпителием среди тканей данного типа является мезотелий. Он покрывает серозные оболочки — листки брюшины и плевры, эпикард и перикард.
Мезотелий — однослойный плоский эпителий, состоящий из плоских эпителиоцитов (мезотелиоцитов). При световой микроскопии плоскостных препаратов видны клеточные границы. Последние хорошо выявляются при импрегнации солями серебра. На первый взгляд клетки мезотелиального пласта мало чем отличаются друг от друга. Однако методами гисторадиоавтографии показано, что в составе дифферона плоских эпителиоцитов имеются различно дифференцированные клетки, что определяется термином гетероморфия. Есть эпителиоциты, делящиеся митозом (камбиальные), дву-ядерные и многоядерные клетки, есть и гибнущие клетки. Между мезотелиоцита-ми имеются контакты типа десмосом. За счет этого мезотелиоциты интегрированы в единую клеточную систему, имеющую вид пласта. Вместе с тем мезотелий — в связи с его положением в своеобразной внутренней изофизиологической среде — утратил некоторые свойства пограничных тканей.
Главные функции мезотелия — покровная, разграничительная, секреторная. Благодаря последней мезотелиальная выстилка создает необходимые условия для скольжения соприкасающихся органов (например, органов брюшной полости) друг относительно друга. Наличие мезотелия на поверхности внутренних органов препятствует образованию спаек, которые ограничивали бы движения органов брюшной полости, легких и сердца.

Физиологическая регенерация мезотелия протекает довольно интенсивно за счет диффузно расположенных в пласте камбиальных эпителиоциов. Для мезотелия характерно слущивание клеток. На их место наползают новые, возникающие в результате деления камбиальных клеток.
К целомическим эпителиям, кроме мезотелия, относятся эпителии органов половой системы — выстилающие извитые семенные канальцы, фолликулярный, эпителий семявыносящих путей, эпителии матки и маточных труб, а также эпителий коры надпочечников. Каждая из этих тканевых разновидностей характеризуется специфическими особенностями детерминации, пролиферации, дифференци-ровки и взаимодействия клеток.
Эпителии нейроглиального типа
Нейроглиальный тип эпителия развивается из нейроэктодермы, выстилает полости мозга и некоторых органов чувств. По строению он однослойный, плоский, кубический или цилиндрический, выполняет вспомогательную для нервных тканей функцию. Иногда этот тип эпи-телиев называют эпендимоглиаль-ным, так как одной из его разновидностей является эпендима, или эпендимный эпителий, образующий выстилку центрального канала спинного мозга и желудочков головного мозга. Эпендимный эпителий — однослойный призматический эпителий. Некоторые авторы рассматривают эпендиму как вспомогательную ткань нервной системы и не относят ее к эпителиям. Кроме эпендимного эпителия среди эпителиев нейроглиального типа различают: эпителий мозговых оболочек, периневральный эпителий, эпителий передней камеры глаза, хрусталиковый эпителий, пигментный эпителий сетчатки глаза, эпителий органа слуха, обонятельный эпителий, вкусовой эпителий, хромаффинный эпителий мозгового вещества надпочечников и адреналовых органов (параганглиев).
Пигментный эпителий сетчатки глаза представляет собой однослойный эпителий, состоящий из клеток полигональной формы. В процессе дифференцировки клетки этой ткани накапливают пигментные включения, необходимые для выполнения эпителием его функции — поглощения световых лучей и изоляции световоспринимающих клеток друг от друга.
Периневральный эпителий — однослойный плоский эпителий, окружающий нервные стволы и выстилающий так называемые периневральные пространства (щели).
Морфофункциональная и гистогенетическая характеристика нейроглиальных эпителиев затруднена недостаточными сведениями в отношении степени их детерминации, особенностей пролиферации и дифференцировки клеток, а также реактивности этих тканей.
Продолжается дискуссия о природе и системной принадлежности сосудистого эндотелия. Н.Г. Хлопин относил эндотелий к эпителиям, выстилающим в виде однослойного пласта внутреннюю поверхность кровеносных, лимфатических сосудов и сердца (эпителий ангиодермального типа). Н.А. Шевченко вьщелил эндотелий в особый тип тканей. СИ. Щелкунов на основе поведения эндотелия в культуре рассматривает сосудистый эндотелий как разновидность тканей внутренней среды. Участие эндотелия гемокапилляров в трансмембранном переносе веществ функционально роднит его с тканями внутренней среды с трофической функцией и поэтому строение эндотелия рассматривается в другом разделе.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Эпителии кожного типа. Эпидермис. Многослойные эпителии.
Эпителии кожного типа развиваются из кожной эктодермы и прехордальной пластинки. Из кожной эктодермы возникают: многослойный плоский ороговевающий эпителий кожи (эпидермис), многослойный плоский неороговевающий эпителий роговицы, эпителий преддверия ротовой полости, эпителии слюнных, потовых, сальных и молочных желез, переходный эпителий мочевыводящих путей и др.
Из прехордальной пластинки развиваются многослойный плоский неороговевающий эпителий пищевода, многорядный мерцательный эпителий воздухоносных путей, однослойный альвеолярный эпителий легких, эпителий щитовидной, околощитовидной, вилочковой желез и передней доли гипофиза.
По своему строению эпителии кожного типа могут быть многослойные, многорядные и однослойные. Многослойные эпителии состоят из нескольких клеточных слоев, из которых лишь базальный слой прилежит к базальной мембране. Клетки базального слоя — эпителиоциты — способны интенсивно делиться митозом. Они служат источником пополнения клеточного состава вышележащих слоев. Базальные эпителиоциты имеют призматическую форму. По мере смещения этих клеток в поверхностные слои они постепенно уплощаются. В многослойном плоском ороговевающем эпителии поверхностный слой образуют роговые чешуйки.
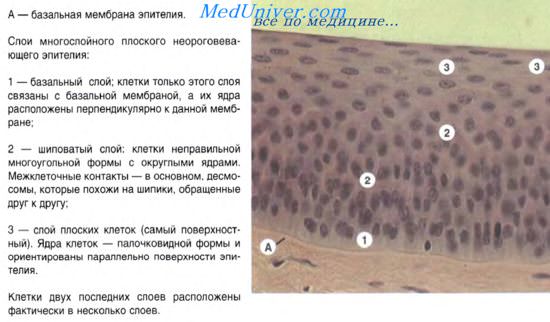
Пограничное положение большинства эпителиев обусловливает определенную цитоархитектонику ткани, а также специфические особенности внутренней структуры клеток и их объединения за счет формирования различных типов межклеточных контактов.
Эпидермис является наиболее типичной разновидностью среди покровных эпителиев. Это полидифферонная ткань. Эпителиальный дифферон развивается из материала кожной эктодермы, отличается стойкой детерминированностью. Диффероны меланоцитов, клеток Лангерганса и клеток Меркеля развиваются из иных источников. Эпителиальный дифферон формирует многослойный пласт ороговевающих клеток (многослойный плоский ороговевающий эпителий). В нем различают слои: базальный, шиповатый, зернистый и роговой. В базальном слое находятся малодифференцированные клетки (базальные эпителиоциты) призматической формы, которые путем митотического деления обеспечивают обновление клеточного состава ткани. После митоза эти клетки перемещаются в вышележащий — шиповатый — слой, образуя клетки многоугольной формы. Клетки шиповатого слоя (шиповатые, крылатые, или остистые, эпителиоциты) имеют в цитоплазме специализированные структуры — тонофиламенты. При световой микроскопии агрегаты тонофиламентов описывают как тонофибриллы. За счет опорных свойств последних достигается механическая прочность клеточного пласта. Между клетками образуются связующие комплексы, или межклеточные контакты — десмосомы.
Следующую стадию дифференцировки составляют уплощенные эпителиоциты зернистого слоя. В цитоплазме этих клеток кроме тонофиламентов синтезируются и накапливаются белки — филаггрин и кератолинин. Ядра зернистых клеток постепенно пикнотизируются, органеллы распадаются под влиянием внутриклеточных ферментов.
Блестящий слой хорошо выявляется только в эпидермисе ладоней и подошв при световой микроскопии. Его образуют плоские постклеточные структуры — кератиноциты, в которых ядра и органеллы исчезают. Из последних образуются роговые чешуйки поверхностного слоя. Они имеют вид 14-гранника. Между чешуйками находится цемонтирующее вещество, богатое липидами (церамиды и др.). Роговые чешуйки имеют плотную оболочку (толщиной 15 им), образованную кератолинином (инволюкрином), ковалентно связанным с оболочкой чешуйки. Содержимое чешуйки заполнено фибриллами зрелого кератина, который характеризуется водонерастворимостью и высокой стойкостью к химическим агентам. Созревание кератина — это агрегация филаментов и обогащение серой за счет образования внутримолекулярных поперечных дисульфидных связей. Этот процесс инициируется филаггрином и происходит при переходе эпителиоцитов из зернистого слоя в роговой. Самые поверхностные слои чешуек постепенно утрачивают связи друг с другом и слущиваются.
Разновидностями многослойных эпителиев являются кубические и призматические эпителии, например, выводных протоков слюнных желез и некоторых других органов, а также многослойный плоский неороговеващий эпителий роговицы. Последний состоит из базального, шиповатого и слоя плоских эпителиоцитов.
Особый вид — переходный эпителий мочевыводящих путей. Он образован базальным, промежуточным и поверхностным слоями. Базальный (камбиальный) слой образован мелкими эпителиоцитами. Полигональные эпителиоциты располагаются в промежуточном слое, а крупные — 2-3-ядерные эпителиоциты — в поверхностном слое. При растяжении мочевого пузыря его стенка уплощается и эпителий растягивается, становится тонким, двуслойным и наоборот, при сокращении — эпителий утолщается. Эпителиоциты промежуточного слоя, не теряя связи с базальной мембраной, становятся грушевидными, а поверхностные — куполообразные.
Многорядный эпителий (ложномногослойный) содержит клетки разной формы. Производными эпителиального дифферона являются реснитчатые, вставочные эпителиоциты, бокаловидные экзокриноциты и эндокриноциты. Располагаются все клетки на базальной мембране. Но вследствие разной высоты ядра эпителиоциов находятся на разных уровнях, что создает впечатление многослойности.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Гистогенез кожи. Строение кожи. Эпидермис. Меланоциты.
В коже как в органе выделяют два слоя: наружный, который образован многослойным плоским ороговевающим эпителием — эпидермисом, и внутренний, состоящий из волокнистой соединительной ткани. Последний именуется собственно дермой. В эпидермисе ведущим клеточным диффероном является эпителиальный, который развивается из кожной эктодермы. Источником развития тканей дермы служит мезенхима дерматомов сомитов.
В первые недели эмбриогенеза эпителий кожи состоит из одного слоя плоских клеток, к концу 2-го месяца становится двухслойным, а на 3-м месяце — многослойным. На 3-м месяце эмбриогенеза вследствие усложнения эпндермо-дермальных взаимодействий в коже появляются базальная мембрана, зачатки желез, волос, ногтей. Параллельно с развитием эпидермиса осуществляются гистогенетические процессы в соединительной ткани кожи, формируются слои дермы (сосочковый и сетчатый), возникает подкожная жировая клетчатка (гиподерма).
В течение 3-го и 4-го месяцев на ладонях и подошвах, включая пальцы, появляются поверхностные гребешки и бороздки. Характер возникающих при этом узоров имеет наследственную природу и не меняется в течение жизни человека. Это используется в методе дактилоскопии, применяемом в криминалистике. На 3-4-м месяцах в эпидермис проникают пигментные клетки — меланобласты и меланоциты, происходящие из материала нервного гребня, затем производные стволовой кроветворной клетки — клетки Лангерганса, а на 5-м месяце — подрастают нервные окончания чувствительных нейронов, позиционную информацию которым передают клетки Меркеля — нейроэндокринные клетки APUD-серии.
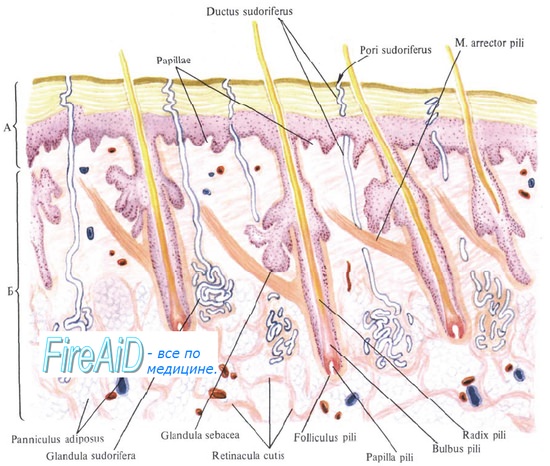
Гисто- и органогенез кожи и ее производных характеризуется индуктивными взаимодействиями эпидермиса и дермы. Вначале в качестве индуктора выступает мезенхима, затем усиливается роль эпидермиса, приводящая к значительному усложнению структуры и функции кожи как органа.
Строение кожи.
Эпидермис — полидифферонная ткань. Эпителиоцнты формируют многослойный ороговевающий пласт клеток, в котором различают два основных функциональных слоя — ростковый и роговой. Эти слои подразделяются на большее количество клеточных слоев в зависимости от толщины эпидермиса и других гистотопографических особенностей кожи.
В наиболее толстом эпидермисе ладоней и подошв, испытывающем сильные воздействия внешних факторов, насчитывается 5 слоев клеток: базальный, шиповатый, зернистый, блестящий (выявляется только при световой микроскопии) и роговой. В составе тонкой кожи блестящий слой отсутствует.
В эпидермисе присутствуют 4 клеточных дифферона: эпителиоциты, меланоциты, клетки Лангерганса и клетки Меркеля. Около 85% от общего числа клеток составляют эпителиоциты, или кератиноциты. Подразделение эпидермиса на слои обусловлено вертикально направленной дифференцировкой эпителиоцитов в составе эпидермально-пролиферативной единицы — гистиона эпидермиса. Последний представлен колонкой эпителиоцитов всех слоев эпидермиса, возникших в результате пролиферации и дифференцировки одной стволовой клетки.
Пространственно гистион имеет вид шестиугольной колонки и по площади равен одной роговой чешуйке. В гистионе также присутствует клетка Лангерганса. Эпителиальные клетки гистиона имеют рецепторы к ряду факторов, регулирующих их развитие — эпидермальному фактору роста и фактору роста кератиноцитов, стимулирующим пролиферацию, к кейлонам, тормозящим пролиферацию эпителиоцитов.
Меланоциты — отростчатые пигментные клетки нейроглиальной природы, расположенные в базальном слое эпидермиса. Их предшественниками являются клетки меланобласты, мигрирующие в состав эпителия из нервного гребня и мозаично встраивающиеся среди эпителиоцитов. Количество меланоцитов может достигать 10% и более от общего числа клеток базального слоя. В их цитоплазме выявляются гранулы темно-коричневого пигмента — меланина, а цитохимическим маркером меланоцитов служит фермент тирозиназа.
Синтез меланина происходит в специальных мембранных органеллах — меланосомах. Кожный пигмент из меланоцитов выделяется в межклеточное пространство и эндоцитозом поступает в эпителиоциты, концентрируясь в зоне ядер и защищая клетки от мутагенного влияния ультрафиолетовых лучей. Меланин может также передаваться по отросткам в клетки подлежащей соединительной ткани дермы — в меланодесмоциты, или меланофоры. Меланин существует в двух формах — эумеланина (черного) и феомеланина (красного). Первый является фотопротектором. Феомеланин не обладает таким свойством и преобладает в коже рыжеволосых людей, которые чувствительны к УФ облучению.
Количество пигмента меняется в зависимости от многих внешних и внутренних факторов. Сильная защитная пигментация кожи (например при загаре) развивается при действии ультрафиолетовых лучей. Пигментация наблюдается также при беременности. В условиях патологии из меланоцитов образуются злокачественные опухоли — меланомы.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

Кожа состоит из трех слоев: эпидермиса, дермы (собственно кожи) и гиподермы (подкожной клетчатки).
Эпидермис – самый верхний, наружный слой кожи - состоит из клеток многослойного плоского ороговевающего эпителия, которые по мере дифференцировки продвигаются от базальной мембраны по направлению к поверхности кожи. Эпидермис, в свою очередь, состоит из 5 слоев: базального, шиповатого, зернистого, блестящего и рогового.
Основа эпидермиса – самый нижний, внутренний базальный слой (stratum basale, germinativum), состоит из 1 ряда мелких клеток цилиндрической формы - базальных кератиноцитов. Непосредственно над базальным слоем кератиноциты увеличиваются в размере и формируют шиповатый слой (stratum spinosum), состоящий из 3—6 (до 15) рядов шиповатых кератиноцитов, постепенно уплощающихся к поверхности кожи. Из-за высокой митотической активности (частого деления клеток) базальный и шиповатый слои называют ростковым слоем Мальпиги, за счет него происходят формирование и регенерация эпидермиса.
Зернистый слой (stratum granulosum) состоит из 2—3 рядов клеток, имеющих вблизи шиповатого слоя цилиндрическую или кубическую форму, а ближе к поверхности кожи – ромбовидную.
Блестящий слой (stratum lucidum) выражен в участках наиболее развитого эпидермиса, т. е. на ладонях и подошвах, где состоит из 3—4 рядов вытянутых по форме слабо контурированных клеток.
Роговой слой (stratum corneum) образован полностью ороговевшими безъядерными клетками – корнеоцитами (роговыми пластинками), которые содержат нерастворимый белок - кератин. Наиболее развит роговой слой там, где кожа подвергается наибольшему механическому воздействию (ладони, подошвы).
Кроме вышеперечисленных клеток в эпидермисе также обнаруживаются меланоциты (клетки, вырабатывающие основной пигмент кожи), а также клетки иммунной системы кожи: клетки Лангерганса (эпидермальные макрофаги) и клетки Гринстейна (тканевые макрофаги).
Эпидермис отделен от дермы базальной мембраной, которая является эластической опорой, прочно связывающей эпителий с дермой и препятствующей прорастанию клеток эпидермиса в дерму.

Дерма – соединительнотканная часть кожи – является опорой для придатков кожи (волос, ногтей, потовых и сальных желез), сосудов и нервов. Она состоит из двух слоев: сосочкового и сетчатого.
Тонкий верхний сосочковый слой (stratum papillare) состоит из бесструктурного вещества и тонких соединительнотканных (коллагеновых, эластических и ретикулярных) волокон и образует сосочки, залегающие между эпителиальными гребнями шиповатых клеток. Более толстый сетчатый слой (stratum reticulare) распространяется от основания сосочкового слоя до подкожной жировой клетчатки; основу его составляют пучки толстых коллагеновых волокон, расположенные параллельно поверхности кожи. В дерме располагается поверхностная сосудистая сеть, и две сети лимфатических сосудов - поверхностная и глубокая. В дерме же сосредоточена основная часть нервного аппарата кожи: два нервных сплетения - глубокое и поверхностное и множественные кожные рецепторы (свободные и инкапсулированные: колбы Краузе,тельца Фатера—Пачини, тельца Мейснера, диски Меркеля). В некоторых участках сосочкового слоя дермы располагаются мышечные волокна, связанные с волосяными луковицами - мышцы, поднимающие волос.
Гиподерма – подкожная жировая клетчатка - состоит из сети коллагеновых, эластических и ретикулярных волокон, между которыми располагаются дольки жировой ткани – скопления крупных жировых клеток-липоцитов. Толщина гиподермы варьирует от 2 мм (на черепе) до 10 см и более (на ягодицах). Гиподерма толще на дорсальных и разгибательных, тоньше на вентральных и сгибательных поверхностях конечностей. Местами (на веках, под ногтевыми пластинками, на крайней плоти, малых половых губах и мошонке) она отсутствует. В подкожной жировой клетчатке залегают крупные сосуды и нервные стволы.
Придатки кожи
К придаткам кожи относят волосы, ногти, сальные и потовые железы.
Волосы. В каждом волосе (pilus) различают две части: стержень и корень. Стержень – часть волоса, выступающая над поверхностью кожи. Корень волоса заложен в дерме и иногда доходит до подкожной жировой клетчатки. Корень волоса погружен в дермальное влагалище (соединительнотканную сумку) и формирует вместе с ним волосяной фолликул. Волосяной фолликул имеет цилиндрическую форму и открывается на поверхности кожи своеобразным расширением – воронкой, в которую погружается стержень волоса. На границе верхней и средней трети фолликула в него открывается выводной проток сальной железы. Самая глубокая расширенная часть корня волоса называется волосяной луковицей; нижняя часть луковицы – матрикс – состоит из клеток, отличающихся очень высокой митотической активностью и обеспечивающих рост волоса.
Анатомия и физиология кожи
Кожа – наш самый большой орган, составляющий 15% от общей массы тела. Она выполняет множество функций, прежде всего защищает организм от воздействия внешних факторов физической, химической и биологической природы, от потери воды, участвует в терморегуляции. Последние научные данные подтверждают, что кожа не только обладает собственной иммунной системой, но и сама является периферическим иммунном органом.
Структура кожи
Кожа состоит из 3 слоев: эпидермиса, дермы и подкожной жировой клетчатки (ПЖК) (рис. 1). Эпидермис – самый тонкий из них, представляет собой многослойный ороговевающий эпителий. Дерма – средний слой кожи. Главным образом состоит из фибрилл структурного белка коллагена. ПЖК содержит жировые клетки – адипоциты. Толщина этих слоев может значительно варьировать в зависимости от анатомического места расположения.
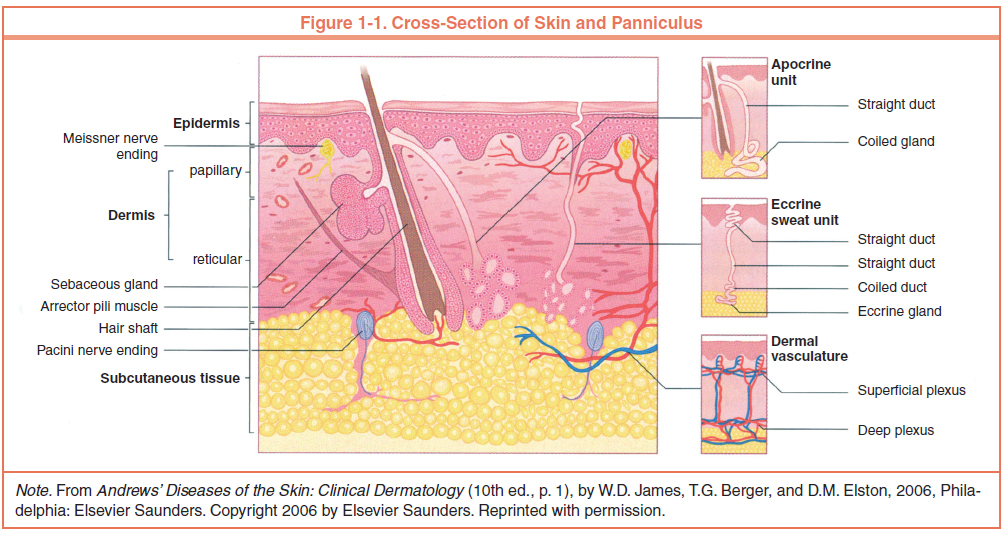
Рис.1. Структура кожи
Эпидермис
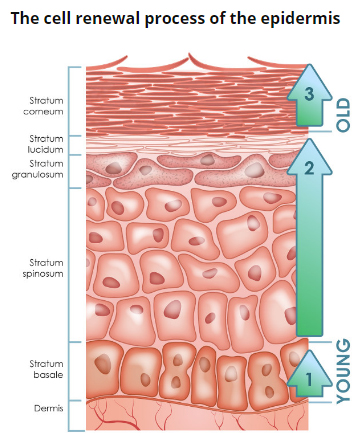
Рис. 2. Эпидермис
Эпидермис – постоянно слущивающийся эпителиальный слой кожи, в котором представлены в основном из 2 типа клеток – кератиноциты и дендритные клетки. В небольшом количестве в эпидермисе присутствуют меланоциты, клетки Лангерганса, клетки Меркеля, внутриэпидермальные Т-лимфоциты. Структурно эпидермис разделяется на 5 слоев: базальный, шиповатый, зернистый, блестящий и роговой , различающиеся положением и степенью дифференцировки кератиноцитов, основной клеточной популяции эпидермиса (рис. 2).
Кератинизация. По мере дифференцировки кератиноцитов и продвижения от базального слоя до рогового происходит их кератинизация (ороговевание) – процесс, начинающийся с фазы синтеза кератина кератиноцитами и заканчивающийся их клеточной деградацией. Кератин служит строительным блоком для промежуточных филаментов. Пучки из этих филаментов, достигая цитоплазматический мембраны, формируют десмосомы, необходимые для образования прочных контактов между соседними клетками. Далее, по мере процесса эпителиальной дифференцировки, клетки эпидермиса вступают в фазу деградации. Ядра и цитоплазматические органеллы разрушаются и исчезают, обмен веществ прекращается, и наступаетапоптозклетки, когда она полностью кератинизируется (превращается в роговую чешуйку).
Базальный слой эпидермиса состоит из одного ряда митотически активных кератиноцитов, которые делятся в среднем каждые 24 часа и дают начало новым клеткам новым клеткам вышележащих эпидермальных слоев. Они активируются только в особых случаях, например при возникновении раны. Далее новая клетка, кератиноцит, выталкивается в шиповатый слой, в котором она проводит до 2 недель, постепенно приближаясь к гранулярному слою. Движение клетки до рогового слоя занимает еще 14 дней. Таким образом, время жизни кератиноцита составляет около 28 дней.
Надо заметить, что не все клетки базального слоя делятся с такой скоростью, как кератиноциты. Эпидермальные стволовые клетки в нормальных условиях образуют долгоживущую популяцию с медленным циклом пролиферации.
Шиповатый слой эпидермиса состоит из 5-10 слоев кератиноцитов, различающихся формой, структурой и внутриклеточным содержимым, что определяется положением клетки. Так, ближе к базальному слою, клетки имеют полиэдрическую форму и круглое ядро, но по мере приближения клеток к гранулярному слою они становятся крупнее, приобретают более плоскую форму, в них появляются ламеллярные гранулы, в избытке содержащие различные гидролитические ферменты. Клетки интенсивно синтезируют кератиновые нити, которые, собираясь в промежуточные филаменты, остаются не связанными со стороны ядра, но участвуют в образовании множественных десмосом со стороны мембраны, формируя связи с соседними клетками. Присутствие большого количества десмосом придает этому слою колючий вид, за что он и получил название «шиповатый».
Зернистый слой эпидермиса составляют еще живые кератиноциты, отличающиеся своей уплощенной формой и большим количеством кератогиалиновых гранул. Последние отвечают за синтез и модификацию белков, участвующих в кератинизации. Гранулярный слой является самым кератогенным слоем эпидермиса. Кроме кератогиалиновых гранул кератиноциты этого слоя содержат в большом количестве лизосомальные гранулы. Их ферменты расщепляют клеточные органеллы в процессе перехода кератиноцита в фазу терминальной дифференцировки и последующего апоптоза. Толщина гранулярного слоя может варьировать, ее величина, пропорциональная толщине вышележащего рогового слоя, максимальна в коже ладоней и подошв стоп.
Блестящий слой эпидермиса (назван так за особый блеск при просмотре препаратов кожи на световом микроскопе) тонкий, состоит из плоских кератиноцитов, в которых полностью разрушены ядра и органеллы. Клетки наполнены элейдином – промежуточной формой кератина. Хорошо развит лишь на некоторых участках тела – на ладонях и подошвах.
Роговой слой эпидермиса представлен корнеоцитами (мертвыми, терминально-дифференцированными кератиноцитами) с высоким содержанием белка. Клетки окружены водонепроницаемым липидным матриксом, компоненты которого содержат соединения, необходимые для отшелушивания рогового слоя (рис. 3). Физические и биохимические свойства клеток в роговом слое различаются в зависимости от положения клетки внутри слоя, направляя процесс отшелушивания наружу. Например, клетки в средних слоях рогового слоя обладают более сильными водосвязывающими свойствами за счет высокой концентрации свободных аминокислот в их цитоплазме.
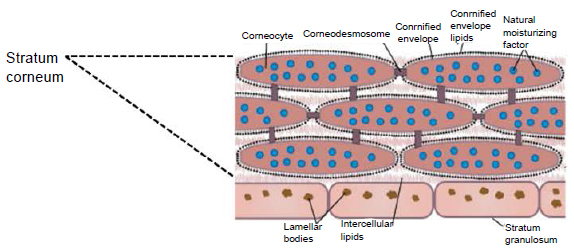
Рис. 3. Схематичное изображение рогового слоя с нижележащим зернистым слоем эпидермиса.
Регуляция пролиферации и дифференцировки кератиноцитов эпидермиса . Являясь непрерывно обновляющейся тканью, эпидермис содержит относительно постоянное число клеток и регулирует все взаимодействия и контакты между ними: адгезию между кератиноцитами, взаимодействие между кератиноцитами и мигрирующими клетками, адгезию с базальной мембраной и подлежащей дермой, процесс терминальной дифференцировки в корнеоциты. Основной механизм регуляции гомеостаза в эпидермисе поддерживается рядом сигнальных молекул – гормонами, факторами роста и цитокинами. Кроме этого, эпидермальный морфогенез и дифференцировка частично регулируются подлежащей дермой, которая играет критическую роль в поддержании постнатальной структуры и функции кожи.
Дерма
Дерма представляет собой сложноорганизованную рыхлую соединительную ткань, состоящую из отдельных волокон, клеток, сети сосудов и нервных окончаний, а также эпидермальных выростов, окружающих волосяные фолликулы и сальные железы. Клеточные элементы дермы представлены фибробластами, макрофагами и тучными клетками. Лимфоциты, лейкоциты и другие клетки способны мигрировать в дерму в ответ на различные стимулы.
Дерма, составляя основной объем кожи, выполняет преимущественно трофическую и опорную функции, обеспечивая коже такие механические свойства, как пластичность, эластичность и прочность, необходимые ей для защиты внутренних органов тела от механических повреждений. Также дерма удерживает воду, участвует в терморегуляции и содержит механорецепторы. И, наконец, ее взаимодействие с эпидермисом поддерживает нормальное функционирование этих слоев кожи.
В дерме нет такого направленного и структурированного процесса клеточной дифференцировки, как в эпидермисе, тем не менее в ней также прослеживается четкая структурная организация элементов в зависимости от глубины их залегания. И клетки, и внеклеточный матрикс дермы также подвергаются постоянному обновлению и ремоделированию.
Внеклеточный матрикс (ВКМ) дермы , или межклеточное вещество, в состав которого входят различные белки (главным образом коллаген, эластин), гликозаминогликаны, самым известным из которых является гиалуроновая кислота, и протеогликаны (фибронектин, ламинин, декорин, версикан, фибриллин). Все эти вещества секретируются фибробластами дермы. ВКМ представляет собой не беспорядочное скопление всех компонентов, а сложноорганизованную сеть, состав и архитектоника которой определяют такие биомеханические свойства кожи, как жесткость, растяжимость и упругость. К белкам ВКМ прикрепляются кератиноциты эпидермиса, которые тесно состыкованы друг с другом. Именно они и формируют плотный защитный слой кожи. Структура ВКМ также способна оказывать регулирующее влияние на погруженные в него клетки. Регуляция может быть как прямой, так и косвенной. В первом случае белки и гликозаминогликаны ВКМ непосредственно взаимодействуют с рецепторами клеток и инициируют в них специфические пути передачи сигнала. Косвенная регуляция осуществляется посредством действия цитокинов и ростовых факторов, удерживаемых в ячейках сети ВКМ и высвобождаемых в определенный момент для взаимодействия с рецепторами клеток. Структурная сеть ВКМ подвергается ремоделированию ферментами из семейства матриксных металлопротеиназ (ММР). В частности, ММР-1 и ММР-13 инициируют деградацию коллагенов I и III типов. Плотность сети ВКМ дермы неравномерна – в папиллярном слое она более рыхлая, в ретикулярном - значительно плотнее как за счет более близкого расположения волокон структурных белков, так и за счет увеличения диаметра этих волокон.
Коллаген – один из главных компонентов ВКМ дермы. Синтезируется фибробластами. Процесс его биосинтеза сложный и многоступенчатый, в результате которого фибробласт секретирует в экстрацеллюлярное пространство проколлаген, состоящий из трех полипептидных α-цепей, свернутых в одну тройную спираль. Затем мономеры проколлагена ферментивным путем собираются в протяженные фибриллярные структуры различного типа. Всего в коже не менее 15 типов коллагена, в дерме больше всего I, III и V типов этого белка: 88, 10 и 2% соответственно. Коллаген IV типа локализуется в зоне базальной мембраны, а коллаген VII типа, секретируемый кератиноцитами, играет роль адаптерного белка для закрепления фибрилл ВКМ на базальной мембране (рис. 4). Волокна структурных коллагенов I, III и V типов служат каркасом, к которому присоединяются другие белки ВКМ, в частности коллагены XII и XIV типов. Считается, что эти минорные коллагены, а также небольшие протеогликаны (декорин, фибромодулин и люмикан) регулируют формирование структурных коллагеновых волокон, их диаметр и плотность образуемой сети. Взаимодействие олигомерных и полимерных комплексов коллагена с другими белками, полисахаридами ВКМ, разнообразными факторами роста и цитокинами приводит к образованию особой сети, обладающей определенной биологической активностью, стабильностью и биофизическими характеристиками, важными для нормального функционирования кожи. В папиллярном слое дермы волокна коллагена располагаются рыхло и более свободно, тогда как ее ретикулярный слой содержит более крупные тяжи коллагеновых волокон.
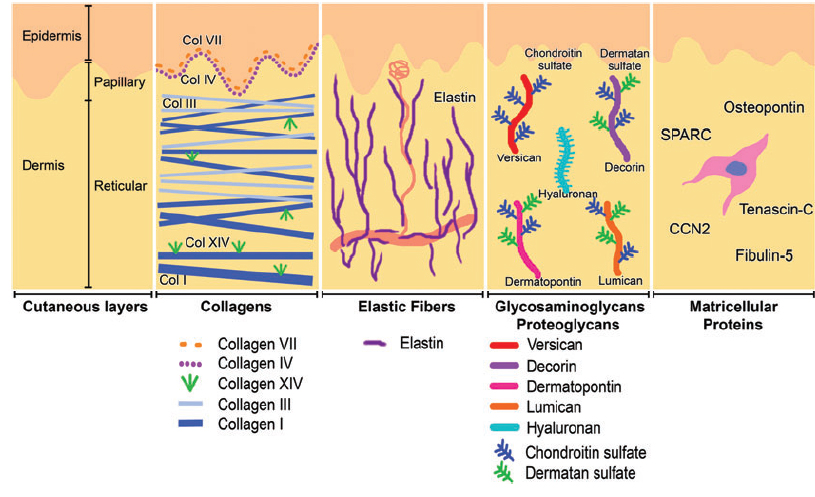
Рис. 4. Схематичное представление слоев кожи и распределения коллагенов разных типов.
Коллаген постоянно обновляется, деградируя под действием протеолитических ферментов коллагеназ и замещаясь вновь синтезированными волокнами. Этот белок составляет 70% сухого веса кожи. Именно коллагеновые волокна «держат удар» при механическом воздействии на нее.
Эластин формирует еще одну сеть волокон в дерме, наделяя кожу такими качествами, как упругость и эластичность. По сравнению с коллагеном эластиновые волокна менее жесткие, они скручиваются вокруг коллагеновых волокон. Именно с эластиновыми волокнами связываются такие белки, как фибулины и фибриллины, с которыми, в свою очередь, связывается латентный TGF-β-связывающий белок (LTBP). Диссоциация этого комплекса приводит к высвобождению и к активации TGF-β, самого мощного из всех факторов роста. Он контролирует экспрессию, отложение и распределение коллагенов и других матриксных белков кожи. Таким образом, интактная сеть из волокон эластина служит депо для TGF-β.
Гиалуроновая кислота (ГК) представляет собой линейный полисахарид, состоящий из повторяющихся димеров D-глюкуроновой кислоты и N-ацетилглюкозамина. Количество димеров в полимере варьирует, что приводит к образованию молекул ГК разного молекулярного веса и длины - 1х10 5 -10 7 Да (2-25 мкм), оказывающих, соответственно, различный биологический эффект.
ГК - высокогидрофильное вещество, влияющее на движение и распределение воды в матриксе дермы. Благодаря этому ее свойству наша кожа в норме и в молодости обладает высоким тургором и сопротивляемостью механическому давлению.
ГК с легкостью образует вторичные водородные связи и внутри одной молекулы, и между соседними молекулами. В первом случае они обеспечивают формирование относительно жестких спиральных структур. Во втором – происходит ассоциация с другими молекулами ГК и неспецифическое взаимодействие с клеточными мембранами, что приводит к образованию сети из полимеров полисахаридов с включенными в нее фибробластами. На длинную молекулу ГК, как на нить, «усаживаются» более короткие молекулы протеогликанов (версикана, люмикана, декорина и др.), формируя агрегаты огромных размеров. Протяженные во всех направлениях, они создают каркас, внося вклад в стабилизацию белковой сети ВКМ и фиксируя фибробласты в определенном окружении матрикса. В совокупности все эти свойства ГК наделяют матрикс определенными химическими характеристиками – вязкостью, плотностью «ячеек» и стабильностью. Однако сеть ВКМ является динамической структурой, зависящей от состояния организма. Например, в условиях воспаления агрегаты ГК с протеогликанами диссоциируют, а образование новых агрегатов между вновь синтезированными молекулами ГК (обновляющимися каждые 3 дня) и протеогликанами блокируется. Это приводит к изменению пространственной структуры матрикса: увеличивается размер его ячеек, меняется распределение всех волокон, структура становится более рыхлой, клетки меняют свою форму и функциональную активность. Все это сказывается на состоянии кожи, приводя к снижению ее тонуса.
Помимо регуляции водного баланса и стабилизации ВКМ, ГК выполняет важную регуляторную роль в поддержании эпидермального и дермального гомеостаза. ГК активно регулирует динамические процессы в эпидермисе, включая пролиферацию и дифференцировку кератиноцитов, окислительный стресс и воспалительный ответ, поддержание эпидермального барьера и заживление раны. В дерме ГК также регулирует активность фибробластов и синтез коллагена. Ремоделируя матрикс, ГК управляет функционированием клеток в матриксе, влияя на их доступность для различных факторов роста и изменяя их функциональную активности. От действия ГК зависит миграция клеток и иммунный ответ в ткани. Таким образом, изменения в распределении, организации, молекулярном весе и метаболизме ГК имеют значимые физиологические последствия.
Фибробласты представляют собой основной тип клеточных элементов дермы. Именно эти клетки отвечают за продукцию ГК, коллагена, эластина, фибронектина и многих других белков межклеточного матрикса, необходимых для формирования соединительной ткани. Фибробласты в различных слоях дермы различаются и морфологически, и функционально. От глубины их залегания в дерме зависит не только количество синтезируемого ими коллагена, но и соотношение типов этого коллагена, например I и III типов, а также синтез коллагеназы: фибробласты более глубоких слоев дермы производят меньшее ее количество. Вообще, фибробласты – очень пластичные клетки, способные менять свои функции и физиологический ответ и даже дифференцироваться в другой тип клеток в зависимости от полученного стимула. В роли последнего могут выступать и сигнальные молекулы, синтезированные соседними клетками, и перестройка окружающего ВКМ.
Подкожно-жировая клетчатка
Подкожно-жировая клетчатка , или гиподерма, - самый нижний слой кожи, располагается под дермой. Состоит из жировых долек, разделенных между собой соединительнотканными септами, содержащими коллаген и пронизанными крупными сосудами. Главными клетками жировых долек являются адипоциты, количество которых варьирует в различных областях тела. В настоящее время ПЖК рассматривают не только как энергетическое депо, но и как эндокринный орган, адипоциты которого участвуют в выработке ряда гормонов (лептина, адипонектина, резистина), цитокинов и медиаторов, оказывающих влияние на метаболизм, чувствительность к инсулину, функциональную активность репродуктивной и иммунной систем.
Читайте также: