
Фильтрация в эпителии это
Обновлено: 26.04.2024
Классификация и отличительные особенности эпителиальных тканей
Эпителиальные ткани (эпителий) – это вид тканей, клетки которых расположены погранично (на границе с внутренней или внешней средой) на базальной мембране в виде пласта тесно, практически без межклеточного вещества, а также образующих большинство желез внешней и внутренней секреции.
Общими характерными особенностями эпителиальных тканей являются:
- погранично расположение;
- клетки различной формы, образующие ряды, слои или пласты;
- полярность клеток;
- небольшое количество межклеточного вещества;
- отсутствие кровеносных, лимфатических сосудов и нервов;
- расположение клеток на базальной мембране, через которую путем диффузии осуществляется газообмен и питание эпителиальных клеток через сосуды, расположенные в подлежащей соединительной ткани;
- высокая способность к регенерации, постоянное обновление;
- способность клеток образовывать реснички и микроворсинки.
В зависимости от выполняемых функций различают две основные группы эпителия: поверхностный (покровный и выстилающий) и железистый.
Функции осморегулирующего и выделительного эпителия
Осморегулирующие и выделительные эпителии относятся к поверхностным, а именно, к выстилающим эпителиям, то есть таким его видам, которые отделяют вторичные полости тела от внутренней среды организма.
Функции осморегуляторного и выделительного эпителия:
- Фильтрационная - выведение из внутренней среды воды, ионов, низкомолекулярных белков, глюкозы и продуктами обмена. Таким образом формируется первичная моча, представляющая собой фильтрат плазмы крови. Суточный объем ее достигает порядка 180 литров, что составляет около 10% от общего объема крови в организме.
- Реабсорбционная - возвращение во внутреннюю среду организма с помощью активного транспорта полезных и нужных ему веществ (глюкозы, аминокислот, небольших белков, почти 80% ионов хлора и натрия). Параллельно в проксимальных отделах канальцев нефронов происходит пассивная реабсорбция воды. За счет реабсорбции из 180 литров первичной мочи образуется до 2 литров вторичной мочи.
- Секреторная - замещение реабсорбируемых ионов натрия ионами калия и аммониевыми ионами, что приводит к закислению мочи. У человека этот процесс осуществляется в дистальном отделе канальца нефрона.
- Накопительная – накопление вторичной мочи в полости мочевого пузыря. Такой процесс стал возможен благодаря переходному эпителию, который по своей сути не является выделительным и осморегулирующим, но часто рассматривается вместе с ними в связи с общностью конечной функции – выделительной.
- Выделительная (экскреторная) - выведение (экскреция) ряда продуктов обмена. Так, почечным эпителием из организма выводятся продукты азотистого обмена и лишняя жидкость. В целом в многоклеточном организме выделение метаболитов его клетками может осуществляться или непосредственно во внешнюю среду, или изначально во внутренние полости организма с дальнейшим выведением наружу.
Готовые работы на аналогичную тему
Строение осморегулирующего и выделительного эпителия
За выполнение каждой отдельной функции отвечает тот или иной вид клеток, каждый из которых приспособлен для выполнения именно этой конкретной задачи.
Эффективным решением функциональной задачи выделения стало возникновение функциональной асимметрии клеток выделительного и осморегулирующего эпителия, что подразумевает непрерывный поток ионов через одну из мембран клетки, где имеются ионные насосы, и постоянное их удаление через другую мембрану, где имеются ионные каналы.
Строение клеток выделительного и осморегулирующего эпителия в связи с их функцией:
Клубочковая ультрафильтрация ( фильтрации ). Регуляция клубочковой фильтрации. Гломерулярный фильтр. Подоциты. Фильтрационное давление.
Процесс клубочковой ультрафильтрации (далее просто фильтрация) осуществляется под влиянием физико-химических и биологических факторов через структуры гломерулярного фильтра, находящегося на пути выхода жидкости из просвета капилляров клубочка в полость капсулы Боумена— Шумлянского.
Гломерулярный фильтр состоит из 3 слоев: эндотелия капилляров, базальной мембраны и эпителия висцерального листка капсулы или подоцитов (см. рис. 14.3). Эндотелий капилляров пронизан отверстиями диаметром до 100 нм. На поверхности эндотелия находится особая выстилка отрицательно заряженными молекулами гликопротеинов, мешающая доступу форменных элементов и крупных молекул, в том числе и белков, к лежащей под эндотелием базальной мембране. Базальная мембрана является основной частью фильтра, препятствующей проникновению из плазмы крови крупномолекулярных соединений (белков). При этом не только размер пор мембраны (около 2,9 нм), но и их отрицательный заряд противодействуют прохождению молекул с отрицательным зарядом, например альбуминов. Базальная мембрана довольно быстро «изнашивается» за счет непрерывного процесса фильтрации, и ее элементы постоянно восстанавливаются с помощью мезангиальных клеток, при этом в течение года происходит полная замена ее основного вещества. Третий слой фильтра образован отростками подоцитов, между которыми остаются щелевые диафрагмы с диаметром пор около 10 нм, поры покрыты гликокаликсом, оставляющим отверстия радиусом около 3 нм. Эта часть фильтра также несет отрицательный заряд.
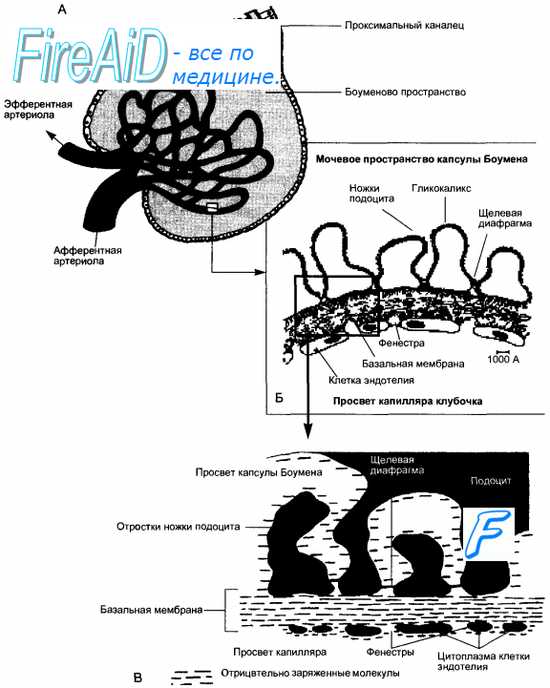
Рис. 14.3. Схема строения клубочка. А — схематическое изображение клубочка в целом, Б — фрагмент трехслойного фильтрационного барьера, В — увеличенный участок фильтрационного барьера. Отчетливо выявляются три слоя барьера: эндотелий капилляра клубочка, базальная мембрана и клетки висцерального листка капсулы Боумена—Шумлянского (подоциты). Фильтрация воды с растворенными в ней веществами происходит из плазмы крови капилляра клубочка через фенестры эндотелия, поры базальной мембраны и щелевые диафрагмы между ножками подоцитов. Все эти структуры фильтрационного барьера имеют отрицательный заряд.
Поскольку подоциты содержат внутри отростков — педикул актомиозиновые миофибриллы, они могут сокращаться и расслабляться, действуя как микронасосы, откачивающие фильтрат в полость капсулы. Эта активность подоцитов составляет один из биологических факторов обеспечения процесса фильтрации, к числу которых относится также сокращение и расслабление мезангиальных клеток, изменяющих тем самым площадь поверхности клубочкового фильтра.
Физико-химические факторы обеспечения фильтрации представлены отрицательным зарядом структур фильтра и фильтрационным давлением, являющимся основной причиной фильтрационного процесса.
Фильтрационное давление — это сила, обеспечивающая движение жидкости с растворенными в ней веществами из плазмы крови капилляров клубочка в просвет капсулы. Эта сила создается гидростатическим давлением крови в капилляре клубочка. Препятствующими фильтрации силами являются онкотическое давление белков плазмы крови (так как белки почти не проходят через фильтр) и давление жидкости (первичной мочи) в полости капсулы клубочка. Таким образом, фильтрационное давление (ФД) представляет собой разность между гидростатическим давлением крови в капиллярах (Рг) и суммой онкотического давления плазмы крови (Ро) и давления первичной мочи (Рм) в капсуле: ФД = Рг — (Ро + Рм). По ходу капилляров клубочка от приносящего к выносящему отделу гидростатическое давление снижается за счет сосудистого сопротивления, а онкотическое давление плазмы, благодаря потере фильтрующейся воды и сгущению, возрастает.
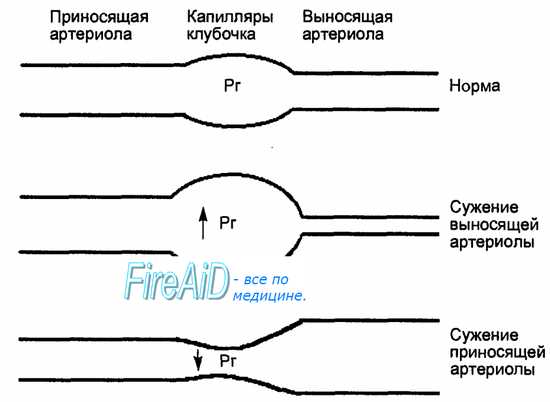
Рис. 14.5. Зависимость величины гидростатического давления в капиллярах клубочка (Рг) от соотношения просветов приносящей и выносящей артериол. При сужении выносящей артериолы гидростатическое давление растет и скорость клубочко-вой фильтрации (СКФ) повышается, а при сужении приносящей артериолы гидростатическое давление и СКФ падают.
Гидростатическое давление крови в приносящей части капилляров клубочка высокое, примерно 50—60 мм рт. ст., т. е. выше, чем в капиллярах других тканей. Это связано, во-первых, с тем, что капилляры клубочка находятся близко к аорте (короткие почечные и внутрипочечные артерии), и, во-вторых,—диаметр приносящих артериол клубочка больше, чем у выносящих.
Гидростатическое давление в выносящей части капилляров ниже на 2—5 мм рт. ст. Гидростатическое давление увеличивается или снижается при изменении соотношения диаметров приносящей и выносящей артериол, что является ведущим механизмом регуляции процесса фильтрации (рис. 14.5). Онкотическое давление белков плазмы крови в приносящей части капилляров клубочка около 25 мм рт. ст., а в выносящей части капилляров, благодаря фильтрации из плазмы воды, оно возрастает до 35—40 мм рт.ст. Давление первичной мочи в капсуле Боумена— Шумлянского примерно равно 15—20 мм рт. ст. Таким образом, ФД в приносящей части капилляров клубочка составляет в среднем: 60 — (25 + 15) = 20 мм рт. ст. В выносящей части капилляров фильтрации практически не происходит, так как ФД равно: 58 — (40 + 15) = 3 мм рт. ст.
Клубочковая фильтрация почек. Состав клубочкового фильтрата
а) Состав клубочкового фильтрата. Образование мочи начинается с фильтрации капиллярами клубочков большого количества жидкости в капсулу Боумена. Как и другие капилляры, сосуды клубочка относительно непроницаемы для белков, поэтому профильтрованная жидкость, называемая первичной мочой, практически не содержит белка, она также свободна от всех клеточных элементов, включая эритроциты.
Концентрации других составляющих первичной мочи, включая большинство солей и органических молекул, схожи с содержанием этих веществ в плазме. Исключениями из правил являются несколько низкомолекулярных веществ, таких как кальций и жирные кислоты, не способные к свободной фильтрации вследствие частичной связи с белками плазмы. Почти половина кальция в плазме и большая часть жирных кислот связаны с белками, поэтому эти вещества не проходят через почечный фильтр.
б) СКФ составляет около 20% почечного плазмотока. Как и в других капиллярах, СКФ определяется: (1) соотношением между гидростатическим и онкотическим давлениями, действующими через стенку капилляра; (2) коэффициентом фильтрации в капилляре (Кф), определяемым проницаемостью мембраны и площадью поверхности капилляров. Капилляры клубочка отличает значительно больший, по сравнению с другими, объем фильтрации вследствие высокого уровня гидростатического давления и значительного Кф В среднем СКФ у взрослого человека составляет 125 мл/мин или 180 л/сут. Часть плазмы, которая подвергается фильтрации в почках (фракция фильтрации), составляет величину 0,2. Это означает, что 20% плазмы, проходящей через почки, фильтруется в клубочках.
Фракцию фильтрации рассчитывают по формуле: Фракция фильтрации = СКФ/Объем плазмотока через почку.
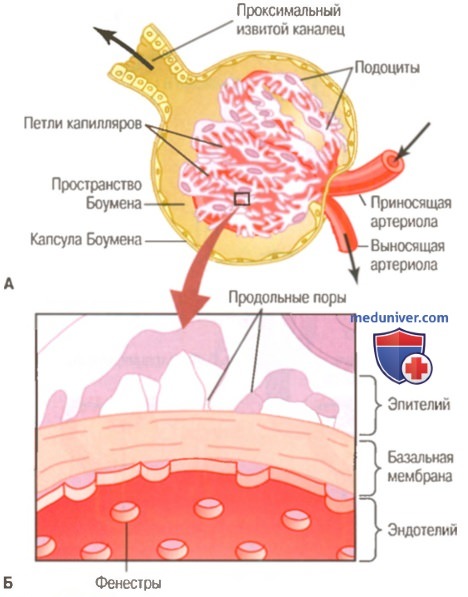
А. Ультраструктура капилляра почечного клубочка.
Б. Поперечное сечение мембраны почечного клубочка и его основных компонентов: эндотелия капилляра, базальной мембраны и эпителия (подоцитов)
в) Мембрана капилляров клубочков — трехслойная (у других капилляров — двуслойная) и состоит из: (1) эндотелия капилляра; (2) базальной мембраны; (3) слоя эпителиальных клеток (подоцитов), окружающих наружную поверхность базальной мембраны капилляров. Эти слои, соединенные вместе, создают фильтрационный барьер, который, несмотря на трехслойность, способен пропускать в сотни раз больше воды и растворенных веществ, чем в обычном капилляре. Даже при высоких темпах фильтрации мембрана клубочков непроницаема для белков.
Высокие параметры фильтрации в мембране клубочка отчасти обусловлены ее особенностями. Эндотелий капилляров содержит тысячи небольших отверстий, называемых фенестрами, схожих по строению с фенестрированными капиллярами, обнаруженными в печени. Несмотря на значительный размер фенестр, эти отверстия непроницаемы для белков, т.к. эндо-телиоцитам присущ отрицательный заряд, препятствующий проникновению белков.
Окруженная эндотелием базальная мембрана содержит сеть фибрилл коллагена и протеогликанов, между волокнами сети имеются большие промежутки, через которые могут проникать вода и небольшие молекулы растворенных веществ. Базальная мембрана успешно препятствует фильтрации белков плазмы — отчасти за счет высокого отрицательного заряда, связанного с протеогликанами.
Завершающая часть мембраны клубочка представлена слоем эпителиальных клеток, образующих прерывистую наружную выстилку клубочка. Эти клетки образуют отростки в виде ножек, оплетающих наружную поверхность капилляров. Между ножками расположены промежутки, называемые продольными порами, через которые перемещается первичная моча. Эпителиальные клетки, которые также заряжены отрицательно, дополнительно ограничивают фильтрацию белков. Таким образом, проникновение белков в первичную мочу предотвращается с помощью всех слоев мембраны клубочка.
Видео физиология фильтрации в почках и поддержки осмотического давления крови - профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Петля Генле. Нисходящий ( тонкий ) отдел петли Генле. Дистальный отдел канальцев ( дистальные канальцы ). Собирательные трубки.
Тонкий нисходящий отдел петли Генле, который покрыт плоскими клетками с щелевидными пространствами в цитоплазме шириной до 7 нм, спускающийся в мозговое вещество почки, где поворачивает на 180° и переходит в тонкую восходящую часть, соединяющуюся с дистальным отделом канальцев.
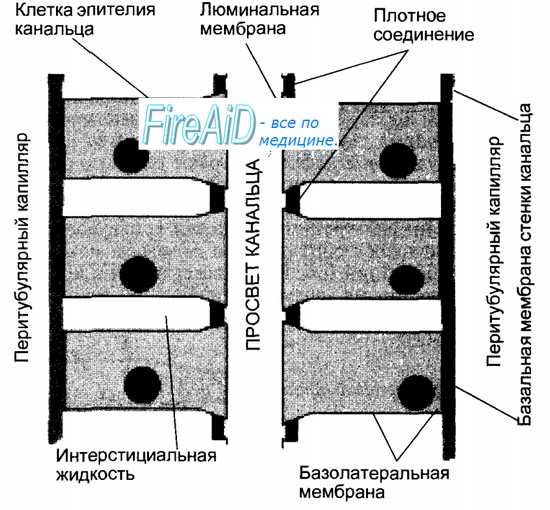
Рис. 14.4. Схема строения почечного канальца. Из просвета канальца через люминальную мембрану клеток эпителия канальца происходит процесс реабсорбции воды и веществ, содержащихся в первичной моче. Поступившие в клетки вещества далее транспортируются через базолатеральные мембраны. Выведение через базо-латеральные мембраны натрия в интерстициальную жидкость способствует повышению ее осмотического давления и обеспечивает осмотический транспорт воды. Из интерстициальной жидкости вода и вещества всасываются в кровь перитубулярных капилляров. Клетки эпителия канальцев обладают также способностью захватывать вещества из кровеносных перитубулярных капилляров и затем секретировать их в просвет канальца через люминальные мембраны.
Дистальный отдел канальцев, состоящий из толстой восходящей части петли Генле или прямого отдела и извитой части. Восходящая часть покрыта клетками, напоминающими клетки проксимального отдела, но лишенными щеточной каемки. Восходящая часть дистального канальца вновь входит в кору почки, подходит к клубочку и обязательно соприкасается с его полюсом между приносящей и выносящей артериолами. Здесь эпителий канальца становится цилиндрическим, ядра клеток — гипер-хромными, этот участок выглядит темным, плотным, что и дало ему название macula densa — плотное пятно. Поскольку непрерывная базальная мембрана здесь отсутствует и клетки эпителия канальца имеют тесный контакт с гранулированными миоэпителиоидными клетками артериолы клубочка (юкстагломерулярными клетками), плотное пятно является структурой юкстагломерулярного аппарата почки. Дистальные извитые канальцы через короткий связующий отдел впадают в коре почек в следующий отдел нефрона — собирательные трубки.
Собирательные трубки спускаются из коры почек вглубь мозгового вещества, где их эпителий из кубического становится цилиндрическим. Темные цилиндрические эпителиальные клетки дистальных отделов собирательных трубок богаты карбоангидразой и обеспечивают секрецию ионов водорода. В глубине мозгового вещества в области вершин пирамид собирательные трубки сливаются в выводные протоки, открывающиеся в полость лоханки.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Цель лекции. Рассмотреть физиологические механизмы всасывания в желудочнокишечном тракте (ЖКТ).
Основные положения. В литературе данные вопросы освещаются с трех сторон: 1) топография всасывания веществ в различных отделах ЖКТ – желудок, двенадцатиперстная кишка, тощая, подвздошная и толстая кишка; 2) основные функции энтероцитов; 3) основные механизмы всасывания в кишечнике. Рассмотрено 7 основных механизмов всасывания веществ в кишечнике.
Заключение. Из всего ЖКТ тощая и подвздошная кишка характеризуются самым широким спектром всасывания различных соединений. Понимание физиологических механизмов всасывания в тонкой кишке имеет большое значение в практической гастроэнтерологии.
Ключевые слова:
Всасывание, ионы, натрий, нутриенты, желудочнокишечный тракт, простая диффузия, облегченная диффузия, осмос, фильтрация, околоклеточный транспорт, активный транспорт, сопряженный транспорт, вторично-энергизованный транспорт, эндоцитоз, трансцитоз, Р-гликопротеин.
![]()
Основные механизмы всасывания

Стенка тонкой кишки, где происходит наиболее интенсивное всасывание основных питательных веществ, или нутриентов, состоит из слизистой оболочки (ворсинки и кишечные железы), подслизистой (где находятся кровеносные и лимфатические сосуды), мышечного слоя (где находятся нервные волокна) и серозной оболочки. Слизистую оболочку образуют ворсинки, покрытые однослойным эпителием с вкраплением бокаловидных клеток; внутри ворсинок проходят лимфатические сосуды, капиллярная сеть, нервные волокна.
Характерная особенность транспорта веществ в эпителии тонкой кишки заключается в том, что он осуществляется через монослой клеток. Всасывающая поверхность такого монослоя существенно увеличена за счет микроворсинок. Энтероциты тонкой кишки, где в основном происходит всасывание питательных веществ (нутриентов), асимметричны, или поляризованы: апикальная и базальная мембраны отличаются друг от друга по проницаемости, набору ферментов, величине разности электрических потенциалов и выполняют неодинаковые транспортные функции.
Ионы попадают в клетки с помощью ионных каналов или специальных молекулярных машин – насосов. Энергия для входа ионов в клетку обычно обеспечивается через плазматическую мембрану электрохимическим градиентом натрия, генерируемым и поддерживаемым благодаря функционированию Na + , K + -АТФазного насоса. Этот насос локализован на базолатеральной мембране, обращенной в кровь (рис. 1).
Энергия, которую можно получить из электрохимического потенциала Na + (разность ионных концентраций + разность электрических потенциалов на мембране) и которая выделяется, когда входящий натрий пересекает плазматическую мембрану, может быть использована другими транспортными системами. Следовательно, Na + , K + -АТФазный насос выполняет две важные функции – откачивает из клеток Na + и генерирует электрохимический градиент, обеспечивающий энергией механизмы входа растворенных веществ.
Термином «всасывание» обозначают совокупность процессов, обеспечивающих перенос веществ из просвета кишки через слой эпителия в кровь и лимфу; секреция – это движение в противоположном направлении.
Всасывание в различных отделах желудочно-кишечного тракта
В желудке всасывается 20% потребленного алкоголя, а также короткоцепочечные жирные кислоты. В двенадцатиперстной кишке – витамины A и B1, железо, кальций, глицерин, жирные кислоты, моноглицериды, аминокислоты, моно- и дисахариды. В тощей кишке – глюкоза, галактоза, аминокислоты и дипептиды, глицерин и жирные кислоты, моно- и диглицериды, медь, цинк, калий, кальций, магний, фосфор, йод, железо, жирорастворимые витамины D, E и K, значительная часть комплекса витаминов В, витамин С и остатки алкоголя. В подвздошной кишке – дисахариды, натрий, калий, хлорид, кальций, магний, фосфор, йод, витамины C, D, E, K, B1, B2, B6, B12 и большая часть воды. В толстой кишке – натрий, калий, вода, газы, некоторые жирные кислоты, образовавшиеся при метаболизме растительных волокон и непереваренного крахмала, витамины, синтезированные бактериями, – биотин (витамин Н) и витамин К.
Основные функции энтероцитов

К основным функциям энтероцитов относят следующие.
Поглощение ионов, включая натрий, кальций, магний и железо, – по механизму их активного транспорта.
Поглощение воды (трансклеточно или околоклеточно), – происходит за счет осмотического градиента, образованного и поддерживаемого ионными насосами, в частности Nа + , К + -АТФазой.
Поглощение сахаров. Ферменты (полисахаридазы и дисахаридазы), локализованные в гликокаликсе, расщепляют большие молекулы сахара на более мелкие, которые затем всасываются. Глюкоза переносится через апикальную мембрану энтероцита с помощью Nа+-зависимого транспортера глюкозы. Глюкоза перемещается через цитозоль (цитоплазму) и выходит из энтероцита через базолатеральную мембрану (в капиллярную систему) с помощью транспортера GLUT-2. Галактоза переносится с помощью такой же транспортной системы. Фруктоза пересекает апикальную мембрану энтероцита, используя транспортер GLUT-5.
Поглощение пептидов и аминокислот. В гликокаликсе ферменты пептидазы расщепляют белки до аминокислот и небольших пептидов. Энтеропептидазы активируют превращение панкреатического трипсиногена в трипсин, который, в свою очередь, активирует другие панкреатические зимогены.
Поглощение липидов. Липиды – триглицериды и фосфолипиды – расщепляются и пассивно диффундируют в энтероциты, а свободные и этерифицированные стерины всасываются в составе смешанных мицелл (см. ниже). Липидные молекулы небольшого размера транспортируются в капилляры кишечника через плотные контакты. Попавшие в энтероцит стерины, включая холестерин, этерифицируются под действием фермента ацил-КоА: холестерин ацилтрансферазы (АХАТ) вместе с ресинтезированными триглицеридами, фосфолипидами и аполипопротеинами включается в состав хиломикронов, которые секретируются в лимфу и затем в кровоток.
Ресорбция неконъюгированных солей желчи. Желчь, попавшая в просвет кишки и не использованная в процессе эмульгации липидов, подвергается обратному всасыванию в подвздошной кишке. Процесс известен как энтерогепатическая циркуляция.
Поглощение витаминов. Для всасывания витаминов используются, как правило, механизмы всасывания других веществ. Особый механизм существует для всасывания витамина В12 (см. ниже).
Секреция иммуноглобулинов. IgA из плазматических клеток слизистой оболочки с помощью механизма рецепторопосредованного эндоцитоза поглощается через базолатеральную поверхность и в виде комплекса рецептор–IgA высвобождается в просвет кишечника. Наличие рецептора придает молекуле дополнительную стабильность.
Основные механизмы всасывания соединений в кишечнике
На рис. 2 представлены основные механизмы всасывания веществ. Рассмотрим указанные механизмы более подробно. Пресистемный метаболизм, или метаболизм (эффект) первого прохождения кишечной стенки. Явление, при котором концентрация вещества перед попаданием в кровеносное русло резко снижается. При этом если введенное вещество является субстратом P-гликопротеина (см. ниже), его молекулы могут неоднократно поступать в энтероциты и выводиться из него, в результате чего вероятность метаболизма данного соединения в энтероцитах возрастает.
P-гликопротеин в большом количестве экспрессирован в нормальных клетках, выстилающих кишечник, проксимальные канальцы почек, капилляры гематоэнцефалического барьера, и в клетках печени. Транспортеры типа P-гликопротеина являются членами надсемейства самого большого и наиболее древнего семейства транспортеров, представленного в организмах от прокариотов до человека. Это трансмембранные белки, функцией которых является транспорт широкого спектра
Пассивный перенос веществ через эпителиальный пласт. Пассивный транспорт веществ через монослой энтероцитов протекает без затрат свободной энергии и может осуществляться или трансклеточным, или околоклеточным путем. К этому виду транспорта относятся простая диффузия (рис. 3), осмос (рис. 4) и фильтрация (рис. 5). Движущей силой диффузии молекул растворенного вещества является его концентрационный градиент.
Зависимость скорости диффузии вещества от его концентрации линейна.Диффузия – это наименее специфичный и самый, по-видимому, медленный процесс транспорта. При осмосе, представляющем собой разновидность диффузионного переноса, происходит перемещение в соответствии с концентрационным градиентом свободных (не связанных с веществом) молекул растворителя (воды).
Околоклеточный транспорт – это транспорт соединений между клетками через область плотных контактов (рис. 7), он не требует затрат энергии. Структура и проницаемость плотных контактов тонкой кишки в настоящее время активно исследуются и дискутируются. Например, известно, что за селективность плотных контактов для натрия отвечает клаудин-2.
Другая возможность состоит в том, что межклеточный перенос осуществляется благодаря некоторым дефектам в эпителиальном пласте. Такое движение может происходить по межклеточным областям в тех местах, где происходит слущивание отдельных клеток.

Такой путь может оказаться воротами для проникновения чужеродных макромолекул прямо в кровь или в тканевые жидкости.
Эндоцитоз, экзоцитоз, рецепторопосредованный транспорт (рис. 8) и трансцитоз. Эндоцитоз – это везикулярный захват жидкости, макромолекул или небольших частиц в клетку. Существуют три механизма эндоцитоза: пиноцитоз (от греческих слов «пить» и «клетка»), фагоцитоз (от греческих слов «поедать» и «клетка») и рецепторопосредованный эндоцитоз или клатрин-зависимый эндоцитоз. Нарушения указанного механизма приводят к развитию определенных заболеваний. Многие кишечные токсины, в частности холерный, попадают в энтероциты именно по этому механизму.
При пиноцитозе гибкая плазматическая мембрана образует впячивание (инвагинация) в виде ямки. Такая ямка заполняется жидкостью из внешней среды. Затем она отшнуровывается от мембраны и в виде везикулы продвигается в цитоплазму, где ее мембранные стенки перевариваются, а содержимое высвобождается. Благодаря такому процессу клетки могут поглощать как крупные молекулы, так и различные ионы, не способные проникнуть через мембрану самостоятельно. Пиноцитоз часто наблюдается в клетках, функция которых связана со всасыванием. Это чрезвычайно интенсивный процесс: в некоторых клетках 100% плазматической мембраны поглощается и восстанавливается всего за час.
При фагоцитозе (явление открыто русским ученым И.И. Мечниковым в 1882 г.) выросты цитоплазмы захватывают капельки жидкости, содержащие какие-либо плотные (живые или неживые) частицы (до 0,5 мкм), и втягивают их в толщу цитоплазмы, где гидролизующие ферменты переваривают поглощенный материал, разрушая его до таких фрагментов, которые могут быть усвоены клеткой. Фагоцитоз осуществляется с помощью клатрин-независимого актин-зависимого механизма; это – основной механизм защиты организма хозяина от микроорганизмов. Фагоцитоз поврежденных или постаревших клеток необходим для обновления тканей и заживления ран.
При рецепторопосредованном эндоцитозе (см. рис. 8) для переноса молекул используются специфические поверхностные рецепторы. Этот механизм обладает следующими свойствами – специфичность, способность к концентрированию лиганда на поверхности клетки, рефрактерность. Если специфический рецептор после связывания лиганда и его поглощения не возвращается на мембрану, клетка становится рефрактерной к данному лиганду.

С помощью эндоцитозного везикулярного механизма всасываются как высокомолекулярные соединения типа витамина В12, ферритина и гемоглобина, так и низкомолекулярные – кальций, железо и др. Роль эндоцитоза особенно велика в раннем постнатальном периоде. У взрослого человека пиноцитозный тип всасывания существенного значения в обеспечении организма питательными веществами, по-видимому, не имеет.
Трансцитоз – это механизм, посредством которого молекулы, пришедшие в клетку извне, могут доставляться к различным компартментам внутри клетки или даже перемещаться от одного слоя клеток к другому. Одним из хорошо изученных примеров трансцитоза является проникновение некоторых материнских иммуноглобулинов через клетки кишечного эпителия новорожденного. Материнские антитела с молоком попадают в организм ребенка. Антитела, связанные с соответствующими рецепторами, сортируются в ранние эндосомы клеток пищеварительного тракта, затем с помощью других пузырьков проходят сквозь эпителиальную клетку и сливаются с плазматической мембраной на базолатеральной поверхности. Здесь лиганды освобождаются от рецепторов. Затем иммуноглобулины собираются в лимфатические сосуды и попадают в кровоток новорожденного.
Рассмотрение механизмов всасывания с точки зрения отдельных групп веществ и соединений будут представлены в одном из следующих номеров журнала.
Работа поддержана грантом РФФИ 09-04-01698
Список литературы:
1. Метельский С.Т. Транспортные процессы и мембранное пищеварение в слизистой оболочке тонкой кишки. Электрофизиологическая модель. – М.: Анахарсис, 2007. – 272 с.
2. Общий курс физиологии человека и животных. – Кн. 2. Физиология висцеральных систем / Под ред. А.Д. Ноздрачева. – М.: Высшая школа, 1991. – С. 356–404.
3. Membrane digestion. New facts and concepts / Ed. A.M. Ugolev. – M.: MIR Publishers, 1989. – 288 p.
4. Tansey T., Christie D.A., Tansey E.M. Intestinal absorption. – London: Wellcome Trust, 2000. – 81 p
статья взята с сайта Русского журнала Гастроэнтерологии, Гепатологии, Колопроктологии
Читайте также: