
Энтодермальный эпителий что это
Обновлено: 25.04.2024
Эпителиальные ткани. Эпителий.
Эпителиальные ткани — древнейшие гистологические структуры, которые в фило- и онтогенезе возникают первыми. Основное свойство эпителиев — пограничность. Эпителиальные ткани (от греч. epi — над и thele — кожица) располагаются на границах двух сред, отделяя организм или органы от окружающей среды.
Эпителии, как правило, имеют вид клеточных пластов и образуют наружный покров тела, выстилку серозных оболочек, просветов органов, сообщающихся с внешней средой во взрослом состоянии или в эмбриогенезе. Через эпителии осуществляется обмен веществ между организмом и окружающей средой. Важной функцией эпителиальных тканей является защита подлежащих тканей организма от механических, физических, химических и других повреждающих воздействий. Некоторые эпителии специализированы на выработке специфических веществ — регуляторов деятельности других тканей организма.
Производными покровных эпителиев являются железистые эпителии. Особый вид эпителия — эпителий органов чувств.
Эпителии развиваются с 3-4-й недели эмбриогенеза человека из материала всех зародышевых листков. Некоторые эпителии, например эпидермис, формируются как полидифферонные ткани, так как в их состав включаются клеточные диффероны, развивающиеся из разных эмбриональных источников (клетки Лангерганса, меланоциты и др.).
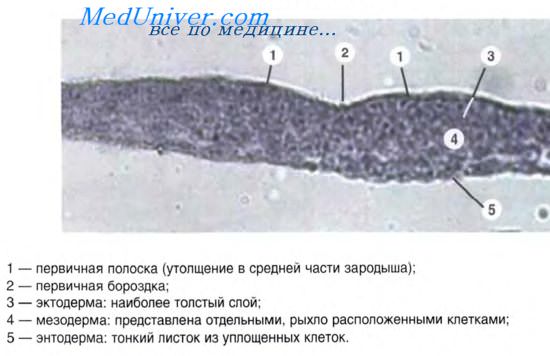
В классификациях эпителия по происхождению за основу, как правило, берется источник развития ведущего клеточного дифферона — дифферона эпителиоцитов. Цитохимическим маркером эпителиоцитов являются белки — цитокератины, образующие тонофиламенты. Цитокератины характеризуются большим разнообразием и служат диагностическим маркером конкретного вида эпителия.
Различают эктодермальные, энтодермальные и мезодермальные эпителии. В зависимости от эмбрионального зачатка, служащего источником развития ведущего клеточного дифферона, эпителии подразделяются на типы: эпидермальный, энтеродермальный, целонефродермальный, эпендимоглиальный и ангиодермальный (Хлопин Н.Г., 1946).
По гистологическим признакам строения ведущего (эпителиального) клеточного дифферона различают однослойные и многослойные эпителии. Однослойные эпителии по форме составляющих их клеток бывают плоские, кубические, призматические или цилиндрические.
Однослойные эпителии подразделяют на однорядные, если ядра всех клеток лежат на одном уровне, и многорядные, в которых ядра расположены на разных уровнях, т. е. в несколько рядов.
Многослойные эпителии подразделяются на ороговевающие и неороговевающие. Многослойные эпителии называют плоскими, учитывая форму клеток наружного слоя. Клетки базального и других слоев могут иметь при этом цилиндрическую или неправильную форму. Кроме названных, выделяют еще переходный эпителий, строение которого меняется в зависимости от степени его растяжения.
На основе данных об органоспецифической детерминации эпителии подразделяются на следующие типы: кожный, кишечный, почечный, целомический и нейроглиальный. В составе каждого типа различают несколько разновидностей эпителиев с учетом их строения и функций. Эпителии перечисленных типов стойко детерминированы. Однако при патологии возможна трансформация одного вида эпителия в другой, но лишь в пределах одного тканевого типа. Например, среди эпителиев кожного типа многорядный мерцательный эпителий воздухоносных путей может переходить в многослойный плоский. Такое явление получило название метаплазия.
Несмотря на разнообразие строения, выполняемых функций и происхождения из разных источников, все эпителии имеют ряд общих признаков, на основе которых их объединяют в систему или группу эпителиальных тканей. Эти общие морфофункциональные признаки эпителиев следующие.
Большинство эпителиев по своей цитоархитектонике представляют собой однослойные или многослойные пласты плотно сомкнутых клеток. Клетки соединены с помощью межклеточных контактов. Эпителий находится в тесных взаимодействиях с подлежащей соединительной тканью. На границе между этими тканями имеется базальная мембрана (пластина). Эта структура участвует в формировании эпителиально-соединительнотканных взаимоотношений, выполняет функции прикрепления с помощью полудесмосом эпителиоцитов, трофическую и барьерную. Толщина базальной мембраны обычно не превышает 1 мкм. Хотя в некоторых органах ее толщина значительно возрастает. Электронно-микроскопически в составе мембраны выделяют светлую (расположенную ближе к эпителию) и темную пластинки. Последняя содержит коллаген IV-ro типа, обеспечивающий механические свойства мембраны. С помощью адгезивных белков — фибронектина и ламинина осуществляется прикрепление эпителиоцитов к мембране. Через базальную мембрану путем диффузии веществ происходит питание эпителия. Базальную мембрану рассматривают в качестве барьера для роста эпителия вглубь. При опухолевых разрастаниях эпителия она разрушается, что позволяет измененным раковым клеткам врастать в подлежащую соединительную ткань (Гаршин В.Г., 1939).
Эпителиоциты обладают гетерополярностью. Строение апикальной и базальной частей клетки разное. В многослойных пластах клетки различных слоев отличаются друг от друга по структуре и функциям. Это называют вертикальной анизоморфией. Эпителии обладают высокой способностью к регенерации за счет митозов камбиальных клеток. В зависимости от местоположения камбиальных клеток в эпителиальных тканях различают диффузный и локализованный камбий.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Эпителии целомического типа. Эпителии нейроглиального типа.
Целомический тип эпителия мезодермалъного происхождения, как правило, однослойный плоский или призматический, выполняет разграничительную, барьерную, секреторную и другие функции. Целомические эпителии развиваются из материала внутренней выстилки сплахнотома, формирующего целом (вторичную полость тела). Наиболее характерным эпителием среди тканей данного типа является мезотелий. Он покрывает серозные оболочки — листки брюшины и плевры, эпикард и перикард.
Мезотелий — однослойный плоский эпителий, состоящий из плоских эпителиоцитов (мезотелиоцитов). При световой микроскопии плоскостных препаратов видны клеточные границы. Последние хорошо выявляются при импрегнации солями серебра. На первый взгляд клетки мезотелиального пласта мало чем отличаются друг от друга. Однако методами гисторадиоавтографии показано, что в составе дифферона плоских эпителиоцитов имеются различно дифференцированные клетки, что определяется термином гетероморфия. Есть эпителиоциты, делящиеся митозом (камбиальные), дву-ядерные и многоядерные клетки, есть и гибнущие клетки. Между мезотелиоцита-ми имеются контакты типа десмосом. За счет этого мезотелиоциты интегрированы в единую клеточную систему, имеющую вид пласта. Вместе с тем мезотелий — в связи с его положением в своеобразной внутренней изофизиологической среде — утратил некоторые свойства пограничных тканей.
Главные функции мезотелия — покровная, разграничительная, секреторная. Благодаря последней мезотелиальная выстилка создает необходимые условия для скольжения соприкасающихся органов (например, органов брюшной полости) друг относительно друга. Наличие мезотелия на поверхности внутренних органов препятствует образованию спаек, которые ограничивали бы движения органов брюшной полости, легких и сердца.

Физиологическая регенерация мезотелия протекает довольно интенсивно за счет диффузно расположенных в пласте камбиальных эпителиоциов. Для мезотелия характерно слущивание клеток. На их место наползают новые, возникающие в результате деления камбиальных клеток.
К целомическим эпителиям, кроме мезотелия, относятся эпителии органов половой системы — выстилающие извитые семенные канальцы, фолликулярный, эпителий семявыносящих путей, эпителии матки и маточных труб, а также эпителий коры надпочечников. Каждая из этих тканевых разновидностей характеризуется специфическими особенностями детерминации, пролиферации, дифференци-ровки и взаимодействия клеток.
Эпителии нейроглиального типа
Нейроглиальный тип эпителия развивается из нейроэктодермы, выстилает полости мозга и некоторых органов чувств. По строению он однослойный, плоский, кубический или цилиндрический, выполняет вспомогательную для нервных тканей функцию. Иногда этот тип эпи-телиев называют эпендимоглиаль-ным, так как одной из его разновидностей является эпендима, или эпендимный эпителий, образующий выстилку центрального канала спинного мозга и желудочков головного мозга. Эпендимный эпителий — однослойный призматический эпителий. Некоторые авторы рассматривают эпендиму как вспомогательную ткань нервной системы и не относят ее к эпителиям. Кроме эпендимного эпителия среди эпителиев нейроглиального типа различают: эпителий мозговых оболочек, периневральный эпителий, эпителий передней камеры глаза, хрусталиковый эпителий, пигментный эпителий сетчатки глаза, эпителий органа слуха, обонятельный эпителий, вкусовой эпителий, хромаффинный эпителий мозгового вещества надпочечников и адреналовых органов (параганглиев).
Пигментный эпителий сетчатки глаза представляет собой однослойный эпителий, состоящий из клеток полигональной формы. В процессе дифференцировки клетки этой ткани накапливают пигментные включения, необходимые для выполнения эпителием его функции — поглощения световых лучей и изоляции световоспринимающих клеток друг от друга.
Периневральный эпителий — однослойный плоский эпителий, окружающий нервные стволы и выстилающий так называемые периневральные пространства (щели).
Морфофункциональная и гистогенетическая характеристика нейроглиальных эпителиев затруднена недостаточными сведениями в отношении степени их детерминации, особенностей пролиферации и дифференцировки клеток, а также реактивности этих тканей.
Продолжается дискуссия о природе и системной принадлежности сосудистого эндотелия. Н.Г. Хлопин относил эндотелий к эпителиям, выстилающим в виде однослойного пласта внутреннюю поверхность кровеносных, лимфатических сосудов и сердца (эпителий ангиодермального типа). Н.А. Шевченко вьщелил эндотелий в особый тип тканей. СИ. Щелкунов на основе поведения эндотелия в культуре рассматривает сосудистый эндотелий как разновидность тканей внутренней среды. Участие эндотелия гемокапилляров в трансмембранном переносе веществ функционально роднит его с тканями внутренней среды с трофической функцией и поэтому строение эндотелия рассматривается в другом разделе.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Эпителиальные ткани. Эпителий.
Эпителиальные ткани — древнейшие гистологические структуры, которые в фило- и онтогенезе возникают первыми. Основное свойство эпителиев — пограничность. Эпителиальные ткани (от греч. epi — над и thele — кожица) располагаются на границах двух сред, отделяя организм или органы от окружающей среды.
Эпителии, как правило, имеют вид клеточных пластов и образуют наружный покров тела, выстилку серозных оболочек, просветов органов, сообщающихся с внешней средой во взрослом состоянии или в эмбриогенезе. Через эпителии осуществляется обмен веществ между организмом и окружающей средой. Важной функцией эпителиальных тканей является защита подлежащих тканей организма от механических, физических, химических и других повреждающих воздействий. Некоторые эпителии специализированы на выработке специфических веществ — регуляторов деятельности других тканей организма.
Производными покровных эпителиев являются железистые эпителии. Особый вид эпителия — эпителий органов чувств.
Эпителии развиваются с 3-4-й недели эмбриогенеза человека из материала всех зародышевых листков. Некоторые эпителии, например эпидермис, формируются как полидифферонные ткани, так как в их состав включаются клеточные диффероны, развивающиеся из разных эмбриональных источников (клетки Лангерганса, меланоциты и др.).
В классификациях эпителия по происхождению за основу, как правило, берется источник развития ведущего клеточного дифферона — дифферона эпителиоцитов. Цитохимическим маркером эпителиоцитов являются белки — цитокератины, образующие тонофиламенты. Цитокератины характеризуются большим разнообразием и служат диагностическим маркером конкретного вида эпителия.
Различают эктодермальные, энтодермальные и мезодермальные эпителии. В зависимости от эмбрионального зачатка, служащего источником развития ведущего клеточного дифферона, эпителии подразделяются на типы: эпидермальный, энтеродермальный, целонефродермальный, эпендимоглиальный и ангиодермальный (Хлопин Н.Г., 1946).
По гистологическим признакам строения ведущего (эпителиального) клеточного дифферона различают однослойные и многослойные эпителии. Однослойные эпителии по форме составляющих их клеток бывают плоские, кубические, призматические или цилиндрические.
Однослойные эпителии подразделяют на однорядные, если ядра всех клеток лежат на одном уровне, и многорядные, в которых ядра расположены на разных уровнях, т. е. в несколько рядов.
Многослойные эпителии подразделяются на ороговевающие и неороговевающие. Многослойные эпителии называют плоскими, учитывая форму клеток наружного слоя. Клетки базального и других слоев могут иметь при этом цилиндрическую или неправильную форму. Кроме названных, выделяют еще переходный эпителий, строение которого меняется в зависимости от степени его растяжения.
На основе данных об органоспецифической детерминации эпителии подразделяются на следующие типы: кожный, кишечный, почечный, целомический и нейроглиальный. В составе каждого типа различают несколько разновидностей эпителиев с учетом их строения и функций. Эпителии перечисленных типов стойко детерминированы. Однако при патологии возможна трансформация одного вида эпителия в другой, но лишь в пределах одного тканевого типа. Например, среди эпителиев кожного типа многорядный мерцательный эпителий воздухоносных путей может переходить в многослойный плоский. Такое явление получило название метаплазия.
Несмотря на разнообразие строения, выполняемых функций и происхождения из разных источников, все эпителии имеют ряд общих признаков, на основе которых их объединяют в систему или группу эпителиальных тканей. Эти общие морфофункциональные признаки эпителиев следующие.
Большинство эпителиев по своей цитоархитектонике представляют собой однослойные или многослойные пласты плотно сомкнутых клеток. Клетки соединены с помощью межклеточных контактов. Эпителий находится в тесных взаимодействиях с подлежащей соединительной тканью. На границе между этими тканями имеется базальная мембрана (пластина). Эта структура участвует в формировании эпителиально-соединительнотканных взаимоотношений, выполняет функции прикрепления с помощью полудесмосом эпителиоцитов, трофическую и барьерную. Толщина базальной мембраны обычно не превышает 1 мкм. Хотя в некоторых органах ее толщина значительно возрастает. Электронно-микроскопически в составе мембраны выделяют светлую (расположенную ближе к эпителию) и темную пластинки. Последняя содержит коллаген IV-ro типа, обеспечивающий механические свойства мембраны. С помощью адгезивных белков — фибронектина и ламинина осуществляется прикрепление эпителиоцитов к мембране. Через базальную мембрану путем диффузии веществ происходит питание эпителия. Базальную мембрану рассматривают в качестве барьера для роста эпителия вглубь. При опухолевых разрастаниях эпителия она разрушается, что позволяет измененным раковым клеткам врастать в подлежащую соединительную ткань (Гаршин В.Г., 1939).
Эпителиоциты обладают гетерополярностью. Строение апикальной и базальной частей клетки разное. В многослойных пластах клетки различных слоев отличаются друг от друга по структуре и функциям. Это называют вертикальной анизоморфией. Эпителии обладают высокой способностью к регенерации за счет митозов камбиальных клеток. В зависимости от местоположения камбиальных клеток в эпителиальных тканях различают диффузный и локализованный камбий.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
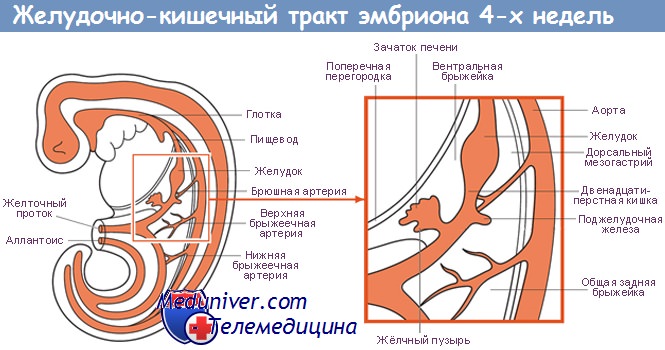
Регуляция специализации энтодермы при формировании желудочно-кишечного тракта у плода
Развитие ЖКТ происходит в три этапа. Первый этап — первичная специализация энтодермы. Второй этап заключается в формировании и структурировании кишечной трубки, что определяет передне-заднюю ось эмбриона и границы между различными органами. На третьем этапе начинается образование органов, которые в процессе развития выходят за границы кишечной трубки, например печени и поджелудочной железы.
В экспериментах на лабораторных животных установлены эволюционно консервативные группы генов, принимающих участие в специализации энтодермы, и ряд других генов, участвующих только в процессе развития ЖКТ позвоночных. Эпителий этого тракта формируется из энтодермы — одного из эмбриональных зародышевых листков, возникающих в процессе гаструляции. Исследования, проведенные на таких лабораторных организмах, как круглые черви (почвенные нематоды Caenorhabdilis elegans), плодовые мушки (род Drosophila), африканские шпорцевые лягушки (род Xenopus), аквариумные рыбки Danio rerio и лабораторные мыши, позволили установить некоторые основные молекулярные механизмы регуляции процесса развития энтодермы. Моделирование на лабораторных мышах, у которых был проведен нокаут определенных генов для исследования их функций, в настоящее время рассматривается в качестве основы для изучения таких заболеваний ЖКТ у человека, как стеноз привратника, различные атрезии, неперфорированный анус.
Специализация энтодермы может быть прослежена на самых ранних стадиях развития эмбриона. В классических экспериментальных исследованиях продемонстрирована способность эксплантатов куриных эмбрионов еще до процесса гаструляции, в культуре, к развитию ЖКТ, что свидетельствует о четкой детерминации процесса уже на данном этапе эмбрионального развития. Энтодерма куриных эмбрионов, выделенная и помещенная в лабораторных условиях в культуру до формирования ЖКТ, характеризуется экспрессией молекулярных маркеров, которые должны в норме появиться in vivo, что свидетельствует о наличии феномена ранней специализации.
Появляются доказательства в поддержку гипотезы, что отличительные особенности энтодермы обусловлены функциональной клеточной независимостью, однако для нормального развития органов необходимо тесное взаимодействие энтодермы и мезодермы. В настоящее время на многочисленных лабораторных организмах установлены целые группы генов, способствующих специализации энтодермы. Один класс генов кодирует факторы транскрипции, которые, в свою очередь, непосредственно активируют гены-мишени. Второй класс генов кодирует синтез сигнальных молекул, обеспечивающих процесс межклеточной кооперации. Как минимум несколько факторов транскрипции, участвующих в процессе специализации энтодермы, продолжают экспрессироваться в тканях ЖКТ в течение всего последующего его развития, например семейства генов Fox и факторов GATA (семейство факторов транскрипции, характеризующихся способностью связываться с фрагментом дезоксирибонуклеиновой кислоты (ДНК) и с GATA-последовательностью нуклеотидов).
В регуляции развития ЖКТ принимают участие, действуя в строго определенное время в различных локусах, сигнальные пути, или механизмы передачи регуляторных стимулов внутри клетки через определенную цепь молекул-посредников, представители группы факторов роста — трансформирующие факторы роста b в том числе непосредственно трансформирующий фактор роста b (TGF-b), и белок морфогенеза костной ткани (BMP), а также сигнальные пути hedgehog (сигнальные системы млекопитающих, представленные тремя различными белками). Продемонстрировано, что фактор транскрипции Sox-17 играет крайне важную роль в развитии энтодермы у мышей на ранних этапах эмбриогенеза, однако роль Sox-17 в дальнейшем не установлена. В последнее время выявлено, что сигнальный путь Notch (рецепторзависимый латентный фактор транскрипции, локализованный в клеточной цитоплазме) также выполняет важную функцию в регуляции развития ЖКТ.
До настоящего времени не удалось выяснить, существует ли «главный ген», запускающий формирование энтодермы и последующее развитие ЖКТ. При исследовании некоторых животных моделей установлены гены, присутствие которых является необходимым и в то же время достаточным условием для формирования энтодермы, например ген mixer у шпорцевых лягушек. Ген мышей mixl1 играет основную роль в дифференцировке эмбриональных клеток в энтодерму. У других лабораторных животных обнаружены гены, необходимые для обеспечения данного процесса, однако недостаточные для формирования полноценной энтодермы.
В исследованиях на лабораторных животных установлены начальные этапы индукции образования энтодермы. Энтодерма у мышей формируется из передней части первичной полоски эпибласта. Ключевой регулятор ранних этапов развития энтодермы — nodal-лиганд факторов роста суперсемейства TGF-b. Nodal экспрессируется в расположенном в бластодерме организационном центре или в узелке первичной бороздки эмбриона мыши. В отсутствие экспрессии nodal эмбрионы мыши не способны к гаструляции или к формированию дифинитивной энтодермы. Аналогично у эмбрионов Danio rerio в отсутствие экспрессии гомологов nodal также не образуется энтодерма. Напротив, экспрессия функционально активных рецепторов к TGF-b способствует преобразованию эмбриональных клеток в энтодерму. Реализация сигнального пути TGF осуществляется посредством стабильного комплекса промежуточных белков Smad.
Необходимым для развития энтодермы является белок Smad2, и нокаут этого белка приводит к нарушению формирования энтодермы задней кишки, а разные мутации в гене, кодирующем Smad2, вызывают различные аномалии передней/головной кишки. FoxA2 также играет важную роль в развитии энтодермы, являясь мишенью сигнального пути, запущенного фактором nodal через белок Smad2. Smad2 и Smad3 совместно регулируют процесс развития энтодермы у мышей, оказывая различное влияние на формирование передней кишки и задней кишки. Кроме того, сигнальная система Smad необходима для развития печени. Основную роль в формировании печени играет ген Hex, экспрессия которого отсутствует либо существенно снижена при нарушенной продукции белков Smad.
Этапы развития желудочно-кишечного тракта у плода
Установлено, что энтодерма находится в тесном контакте с мезодермой на протяжении всего ЖКТ. В экспериментальных исследованиях, выполненных на культурах тканей, было показано, что формирование энтодермы и ее дифференцировка в отдельные органы представляет собой результат взаимодействия мезодермы и энтодермы. Раньше всего было показано, что для формирования переднезадней оси энтодермы мышей необходимо сигнальное воздействие мезодермы на энтодерму посредством фактора роста фибробластов (FGF).
Другие члены семейства FGF и рецепторы к ним необходимы для формирования печени. Еще три семейства генов крайне необходимы в процессе взаимодействия мезодермы и энтодермы — гены, кодирующие факторы сигнальных путей hedgehog и BMP, гены box.
У почвенной нематоды Caenorhabditis elegans для специализации клеток, из которых развивается кишечный эпителий, необходимы два гена, кодирующие факторы транскрипции GATA, в то время как фактор GATA у плодовых мушек рода Drosophila кодируется одним геном serpent, наличие которого нужно для дифференцировки кишечной энтодермы. В кишечнике позвоночных экспрессируются три фактора группы GATA. Предполагается, что в процессах пролиферации и дифференцировки кишечного эпителия факторы GATA4, -5 и -6 выполняют различные функции, однако их функциональное значение на ранних этапах развития кишечной трубки у млекопитающих пока не установлено. Помимо факторов GATA, представители семейства forkhead-related факторов (кодируемые генами Fox) и сигнального пути WNT/Tcf являются важными регуляторами развития энтодермы.
У позвоночных важную роль в инициации формирования энтодермы играют представители суперсемейства TGF-b. Молекула цитоскелета ELF3 задействована в сигнальном пути через факторы TGF-P, а также необходима для развития энтодермы. В отсутствие гена, кодирующего ELF3, у мышей полностью отсутствует кишечная энтодерма.
Многочисленные факторы транскрипции, первоначально считавшиеся специфическими для формирования печени, играют роль и в развитии кишечника. При изучении эмбрионов мыши было отмечено, что некоторые из факторов транскрипции имеют характерные паттерны экспрессии, что позволило предположить их участие в развитии кишечника. Например, нуклеарный печеночный фактор (HNF) 3b (сейчас известный как FoxA2) оказался важным элементом в процессе самых ранних этапов дифференцировки ЖКТ и продолжает экспрессироваться в производных энтодермы у взрослых. Гомозиготные мутанты с полным отсутствием HNF3b не способны к формированию первичной полоски, предшествующей образованию кишечной трубки и ряда других структур. HNF3b участвует в образовании передней и средней, но не задней кишки.
Идентифицировано большое количество факторов семейства Fox, некоторые из которых характеризуются преимущественной или специфической экспрессией в кишечнике. Один из представителей данного семейства, Foxl1, в норме экспрессируется кишечной мезодермой и представляет собой основной медиатор взаимодействия между эпителием и мезенхимой. Отсутствие указанного фактора сопровождается патологической пролиферацией эпителиальных клеток и нарушением развития кишечника. Этот фактор действует через экспрессию протеогликанов, проявляющих себя как корецепторы к компонентам сигнального пути WNT, и таким образом активирует комплекс WNT/P-катенин, регулирующий пролиферацию клеток. Группа исследователей во главе с Zaret предложила модель, в которой факторы FoxA2 и GATA открывают ДНК в клетках энтодермы, обеспечивая ее доступность для последующего связывания с факторами транскрипции, регулирующими клеточно-специфические гены. Таким образом, вероятно, при развитии кишечника многочисленные представители семейства факторов Fox играют важную роль в этом сложном процессе, который окончательно не изучен.
У мышей некоторые гены, гомологичные гену caudal у дрозофил, экспрессируются исключительно в кишечнике. Cdx-1, экспрессирующийся в кишечнике взрослых организмов, также широко экспрессируется в тканях развивающегося эмбриона. Другой ген, Cdx-2, экспрессируется в висцеральной энтодерме на ранних стадиях эмбрионального развития, но впоследствии его паттерн экспрессии ограничен исключительно кишечником. Чрезмерная экспрессия Cdx-2 приводит к дальнейшей дифференцировке популяции клеток кишечника, которые в норме должны оставаться недифференцированными. В то же время эктопическая экспрессия Cdx-2 в стенке желудка приводит к формированию ткани, аналогичной кишечнику. Cdx-2 представляет собой важный фактор дифференцировки кишечника, однако до настоящего времени его роль в формировании кишечной трубки на ранних стадиях эмбрионального развития точно не установлена.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Эпителии целомического типа. Эпителии нейроглиального типа.
Целомический тип эпителия мезодермалъного происхождения, как правило, однослойный плоский или призматический, выполняет разграничительную, барьерную, секреторную и другие функции. Целомические эпителии развиваются из материала внутренней выстилки сплахнотома, формирующего целом (вторичную полость тела). Наиболее характерным эпителием среди тканей данного типа является мезотелий. Он покрывает серозные оболочки — листки брюшины и плевры, эпикард и перикард.
Мезотелий — однослойный плоский эпителий, состоящий из плоских эпителиоцитов (мезотелиоцитов). При световой микроскопии плоскостных препаратов видны клеточные границы. Последние хорошо выявляются при импрегнации солями серебра. На первый взгляд клетки мезотелиального пласта мало чем отличаются друг от друга. Однако методами гисторадиоавтографии показано, что в составе дифферона плоских эпителиоцитов имеются различно дифференцированные клетки, что определяется термином гетероморфия. Есть эпителиоциты, делящиеся митозом (камбиальные), дву-ядерные и многоядерные клетки, есть и гибнущие клетки. Между мезотелиоцита-ми имеются контакты типа десмосом. За счет этого мезотелиоциты интегрированы в единую клеточную систему, имеющую вид пласта. Вместе с тем мезотелий — в связи с его положением в своеобразной внутренней изофизиологической среде — утратил некоторые свойства пограничных тканей.
Главные функции мезотелия — покровная, разграничительная, секреторная. Благодаря последней мезотелиальная выстилка создает необходимые условия для скольжения соприкасающихся органов (например, органов брюшной полости) друг относительно друга. Наличие мезотелия на поверхности внутренних органов препятствует образованию спаек, которые ограничивали бы движения органов брюшной полости, легких и сердца.
Физиологическая регенерация мезотелия протекает довольно интенсивно за счет диффузно расположенных в пласте камбиальных эпителиоциов. Для мезотелия характерно слущивание клеток. На их место наползают новые, возникающие в результате деления камбиальных клеток.
К целомическим эпителиям, кроме мезотелия, относятся эпителии органов половой системы — выстилающие извитые семенные канальцы, фолликулярный, эпителий семявыносящих путей, эпителии матки и маточных труб, а также эпителий коры надпочечников. Каждая из этих тканевых разновидностей характеризуется специфическими особенностями детерминации, пролиферации, дифференци-ровки и взаимодействия клеток.
Эпителии нейроглиального типа
Нейроглиальный тип эпителия развивается из нейроэктодермы, выстилает полости мозга и некоторых органов чувств. По строению он однослойный, плоский, кубический или цилиндрический, выполняет вспомогательную для нервных тканей функцию. Иногда этот тип эпи-телиев называют эпендимоглиаль-ным, так как одной из его разновидностей является эпендима, или эпендимный эпителий, образующий выстилку центрального канала спинного мозга и желудочков головного мозга. Эпендимный эпителий — однослойный призматический эпителий. Некоторые авторы рассматривают эпендиму как вспомогательную ткань нервной системы и не относят ее к эпителиям. Кроме эпендимного эпителия среди эпителиев нейроглиального типа различают: эпителий мозговых оболочек, периневральный эпителий, эпителий передней камеры глаза, хрусталиковый эпителий, пигментный эпителий сетчатки глаза, эпителий органа слуха, обонятельный эпителий, вкусовой эпителий, хромаффинный эпителий мозгового вещества надпочечников и адреналовых органов (параганглиев).
Пигментный эпителий сетчатки глаза представляет собой однослойный эпителий, состоящий из клеток полигональной формы. В процессе дифференцировки клетки этой ткани накапливают пигментные включения, необходимые для выполнения эпителием его функции — поглощения световых лучей и изоляции световоспринимающих клеток друг от друга.
Периневральный эпителий — однослойный плоский эпителий, окружающий нервные стволы и выстилающий так называемые периневральные пространства (щели).
Морфофункциональная и гистогенетическая характеристика нейроглиальных эпителиев затруднена недостаточными сведениями в отношении степени их детерминации, особенностей пролиферации и дифференцировки клеток, а также реактивности этих тканей.
Продолжается дискуссия о природе и системной принадлежности сосудистого эндотелия. Н.Г. Хлопин относил эндотелий к эпителиям, выстилающим в виде однослойного пласта внутреннюю поверхность кровеносных, лимфатических сосудов и сердца (эпителий ангиодермального типа). Н.А. Шевченко вьщелил эндотелий в особый тип тканей. СИ. Щелкунов на основе поведения эндотелия в культуре рассматривает сосудистый эндотелий как разновидность тканей внутренней среды. Участие эндотелия гемокапилляров в трансмембранном переносе веществ функционально роднит его с тканями внутренней среды с трофической функцией и поэтому строение эндотелия рассматривается в другом разделе.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: