
Что такое мембраны клеток кожи
Обновлено: 19.04.2024
Структурное и функциональное здоровье кожи во многом связано с содержанием и распределением в ней воды. Сухость кожи влечет за собой ее легкую проницаемость для токсичных и сенсибилизирующих веществ, способствует развитию иммунных нарушений и формированию
Структурное и функциональное здоровье кожи во многом связано с содержанием и распределением в ней воды. Сухость кожи влечет за собой ее легкую проницаемость для токсичных и сенсибилизирующих веществ, способствует развитию иммунных нарушений и формированию аллергического воспаления [1–3]. Нарушение гидратации рогового слоя эпидермиса является основной причиной патологической десквамации эпителия и ксероза. Поскольку корнеоциты представляют собой клетки, лишенные ядра, дерматологи до последнего времени не относились к роговому слою с должным вниманием. Но оказалось, что, воздействуя на роговой слой, можно не только добиться косметических эффектов, но и облегчить симптомы кожного заболевания, улучшить результат от медикаментозного лечения и повысить качество жизни больного.
Основа ухода за кожей при кожных заболеваниях — «корнеотерапия», система восстановления функционирования и целостности рогового слоя. В современных условиях грань между косметическими и лекарственными средствами постепенно стирается ввиду усложнения рецептуры косметологических препаратов. Современные косметические средства могут влиять на физиологические процессы в коже так же, как это делает фармацевтический препарат. Мастерство дерматолога основано на его умении использовать в терапии различные активные и индифферентные средства наружной терапии, добиваясь максимального успеха.
В поддержании водного баланса участвуют многочисленные структуры, регулирующие скорость трансэпидермальной потери воды и осуществляющие поступление воды из дермы в эпидермис. Очевидно, что для полноценного функционирования кожи необходима опорная структура корнеоцитов, при ее нарушениях увеличивается трансэпидермальная потеря воды, что ведет к гиперплазии эпидермиса и гиперкератозу. Роговой слой служит сенсором для подлежащих слоев [1]. В эпидермисе контроль за водным балансом осуществляет роговой слой и компоненты рогового слоя эпидермиса (натуральный увлажняющий фактор, липиды кожи, кожное сало и кератин) [3].
Кожа начинает обезвоживаться именно с эпидермиса. Происходит это из-за того, что эпидермис лишен кровеносных сосудов, которые могли бы пополнять в нем запасы влаги, поэтому основную часть жидкости эпидермис получает из дермы. До последнего времени считалось, что вода просачивается в верхние слои кожи по закону перфузии. Свободные аминокислоты, образующиеся при разрушении белка филагрина, поддерживают в корнеоцитах высокое осмотическое давление, что вызывает приток воды, которая сохраняется даже при пониженной влажности окружающей среды [4]. Однако в последние годы установлено, что в поддержании нормального уровня гидратации эпидермиса важное значение имеет также механизм облегченной диффузии, т. е. попадание молекул воды через водные каналы, названные аквапоринами. Аквапорины представляют собой мембранные белки, формирующие сквозные поры на поверхности клеток, через которые проходит вода, а также небольшие водорастворимые соединения, такие как глицерин и мочевина. За открытие этих белков в 2003 г. группе американских учёных во главе с Питером Эгром присуждена Нобелевская премия по химии [3].
Достигнув поверхности кожи, вода стремится к испарению, однако этому препятствует роговой слой. Несмотря на низкое содержание воды в роговом слое — 15–20%, сохранение влаги одна из основных его функций [5]. Непроницаемость рогового слоя обусловлена барьерными свойствами кератина, ацилцерамидов и кожного сала. Эпидермис достаточно хорошо проницаем по направлению к своей поверхности и почти непроницаем в обратном направлении. При этом вода и водорастворимые вещества сквозь эпидермис проникают с большим трудом, тогда как жирорастворимые вещества значительно легче [6, 7]. Чтобы оценить важную роль рогового слоя эпидермиса в сохранении влаги в коже, необходимо четко знать его строение.
Роговой слой представляет собой конечный продукт дифференцировки кератиноцитов эпидермиса. В тонкой коже он состоит из 15–20 слоев роговых чешуек, в толстой из сотен слоев. Чешуйки располагаются колонками друг над другом, и каждая чешуйка в проекции покрывает 9–10 клеток базального слоя [8]. Корнеоцит имеет 6-угольную форму и каждой из шести сторон контактирует с соседними корнеоцитами, такое строение можно сравнить со «стеганым одеялом» [4]. Роговой слой эпидермиса имеет уникальное строение, которое называют «briсk and mortar» («кирпич и цемент»), где роль «кирпичей» играют роговые клетки, а «цемента» — межклеточные липиды. На поперечном срезе роговой слой представляется рыхлым, пористым, однако это только видимость. Такое впечатление создается за счет межклеточных пространств, которые составляют значительную часть рогового слоя. Однако все они заполнены особым веществом, склеивающим их в непреодолимую преграду липидами-ацилцерамидами, которые относятся к классу сфинголипидов, или церамидов [6, 7]. Впервые сфинголипиды были выделены из мозговой ткани, поэтому свое второе название — церамиды — они получили от латинского слова cerebrum (мозг). Позже было установлено, что церамиды участвуют в построении эпидермального барьера, формируя липидную прослойку между роговыми чешуйками. Молекулы церамидов и фосфолипидов имеют гидрофильные «головы» (фрагменты, любящие воду) и липофильные «хвосты» (фрагменты, предпочитающие жиры). В водной среде молекулы полярных липидов самостоятельно группируются таким образом, чтобы гидрофобные хвосты были спрятаны от воды, а гидрофильные головы, напротив, были обращены в водную среду. Если таких липидов мало (и если смесь липидов и воды хорошо встряхнуть), то образуются шарики (липосомы). Это свойство полярных липидов используется в косметической промышленности при производстве липосом [5].
Церамиды состоят из жирного спирта сфингозина (образует «голову») и одной жирной кислоты («хвост»). Если в жирной кислоте имеются двойные связи, то она называется ненасыщенной, если двойных связей нет, то говорят, что кислота насыщенная. В зависимости от того, какая жирная кислота прикреплена к голове церамида, липидные пласты, построенные из них, получаются более или менее жидкими. Самые твердые (кристаллические) липидные пласты образованы церамидами с насыщенными хвостами. Чем длиннее хвост церамида и чем больше в нем двойных связей, тем более жидкими получаются липидные структуры [9–11]. Помимо рассмотренных выше межклеточных липидных пластов, в роговом слое обнаружены липиды, ковалентно связанные с корнеоцитами. Это особые длинноцепочечные церамиды, хвосты которых представлены жирными кислотами, имеющими в своей цепочке более 20 атомов углерода. Длинноцепочечные церамиды выполняют роль заклепок, скрепляя соседние липидные пласты. Благодаря им многослойная липидная прослойка не расслаивается и представляет собой целостную структуру. Церамиды в последнее время стали очень популярными ингредиентами в косметике. Популярность церамидов объясняется той ролью, которую они играют в поддержании целостности эпидермального барьера.
Благодаря наличию многослойной липидной прослойки между роговыми чешуйками, роговой слой способен эффективно защищать кожу не только от проникновения посторонних веществ извне, но и от обезвоживания. Диффузия воды через сухие полутвердые кератиновые пластинки, склеенные в сплошную массу церамидами, резко снижается, по сравнению с наполненными жидкостью живыми клетками.
Основными продуцентами межклеточного цемента являются гранулы Орланда клеток зернистого слоя [6, 7]. Именно они путем экзоцитоза выделяют свое содержимое в межклеточное пространство, где оно превращается в богатый липидами межклеточный цемент с пластинчатым строением. Таким образом, при ороговении происходит формирование гидрофобного эпидермального барьера, который не только препятствует проникновению в кожу вредных веществ, но и участвует в регуляции водного гомеостаза в дерме [4].
Кроме механизма биомеханической непроницаемости рогового слоя, влажность кожи поддерживают влагоудерживающие структуры.
Эпидермальные влагоудерживающие структуры кожи
1. Натуральный увлажняющий фактор (natural moisturizing factor, NMF) — это целый комплекс органических молекул на поверхности корнеоцитов, обладающий способностью связывать воду. К нему относятся свободные аминокислоты (40%); пироглутамат натрия (12%); мочевина (7%); аммиак, креатинин и др. органические соединения (17%); магний (1,5%); калий (4%); кальций (1,5%); натрий (5%); молочная и лимонная кислоты, ионы хлорида и фосфата (12%). Нарушение их баланса влечет за собой изменение состава NMF и, как следствие, неспособность кожи удерживать влагу. От количества влаги, связанной с NMF, зависит и эластичность рогового слоя. Доказано, что в жирной и нормальной коже больше NMF, чем в сухой [5].
Снижение синтеза филагрина, которое наблюдается, в частности, при ихтиозе и атопическом дерматите, приводит к уменьшению содержания аминокислот в составе NMF и уменьшению водоудерживающей способности кожи. Так как для работы ферментов, участвующих в гидролизе филагрина, необходимо достаточное количество влаги, при развитии сухости рогового слоя образование аминокислот NMF также снижается. В результате получается замкнутый круг, приводящий к хронической сухости кожного покрова.
Минеральный состав натурального увлажняющего фактора меняется в зависимости от времени года, и именно эти изменения вызывают снижение влажности рогового слоя в зимний период. В частности, в зимний период в роговом слое значительно понижается уровень калия, натрия, хлоридов и солей молочной кислоты [5].
При частом умывании горячей водой с мылом из NMF могут вымываться аминокислоты и минеральные вещества, что тоже приводит к развитию ксероза. Наиболее опасно сочетание «вымачивания», например длительного нахождения в горячей ванне, и воздействия растворителей и агрессивных поверхностно-активных веществ (ПАВ). Опасность ПАВ заключается в том, что они способны взаимодействовать с липидными пленками, поскольку в них, как и в полярных липидах, есть два участка — гидрофобный и гидрофильный, поэтому они могут встраиваться в липидный слой, обращаясь к ним своими гидрофобными «хвостами». При этом головы молекул ПАВ группируются вместе так, что в липидном слое оказываются гидрофильные, проницаемые для воды участки. Таким образом, первым следствием воздействия ПАВ на роговой слой является дегидратация липидных мембран, так называемый «Wash-Out-Effect» — эффект вымывания собственных липидов.
Длительное воздействие анионных ПАВ способствует их более глубокому проникновению вплоть до зернистого слоя эпидермиса, что приводит к разрушению клеточных мембран гранулоцитов — гранул Орланда, являющихся основной фабрикой межклеточных липидов. В результате снижается синтез липидов, необходимых для склеивания корнеоцитов [8].
Именно поэтому любые косметические процедуры, сопровождающиеся «отмачиванием» кожи с последующим ее очищением, должны завершаться применением увлажняющих средств на основе NMF [5, 14].
2. Эпидермальные межклеточные липиды (липидный барьер) — состоят из холестерола, церамидов и ненасыщенных жирных кислот (омега-3, омега-6), количество которых находится в строго определенной пропорции по отношению друг к другу. Если корнеоцит содержит лишь 3% липидов, то межклеточный цемент — 80%. По биохимическим данным в межклеточном матриксе обнаруживаются: церамиды — ~40%, свободные жирные кислоты — ~20%, холестерин и его эфиры — ~10%, холестерол — ~15%, фосфолипиды — ~5%, сквален — ~10% [4]. Холестерин предотвращает избыточную ригидность и ломкость слоя церамидов. Свободные жирные кислоты расположены вокруг церамидов в липидном слое и способствуют поддержанию водоотталкивающей функции кожи, защищают водорастворимые компоненты рогового слоя от выщелачивания вследствие образования водно-масляной эмульсии [4, 9]. Если эта пропорция меняется, липидная прослойка между роговыми чешуйками нарушается и, как следствие, нарушается барьерная функция, влага испаряется более интенсивно. Пытаясь предотвратить избыточную потерю влаги, кожа замедляет процесс физиологической десквамации, и клетки начинают скапливаться на поверхности. Внешне это проявляется избыточным шелушением, утолщением рогового слоя, сероватым оттенком. Ярким примером такого процесса является себорея. При себорее в кожном сале уменьшается концентрация линолевой кислоты, что также приводит к нарушению целостности кожного барьера и возникновению адаптивной реакции в виде увеличения количества роговых клеток [12].
Среди липидов поверхности кожи выявляются эпидермальные липиды и липиды сальных желез. Эпидермальные липиды составляют меньшую часть. Главные компоненты эпидермальных липидов — свободный холестерин и его эфиры — образуются при распаде оболочек клеток рогового слоя. Доказано, что эпидермальные липиды поверхности кожи не оказывают большого влияния на гидратацию кожи, но представляют собой ограничивающую мембрану для водного обмена [4, 6, 7].
Интересно, что липидный состав рогового слоя неоднороден. Концентрация фосфолипидов уменьшается по направлению к поверхности, тогда как содержание нейтральных липидов и церамидов, наоборот, повышается [4].
Изменение состава эпидермальных липидов происходит при многих заболеваниях: атопическом дерматите, псориазе, контактном дерматите, себорее, а также при некоторых физиологических процессах. Например, при атопическом дерматите в коже нарушен обмен жирных кислот, а при ихтиозе наблюдается их снижение (табл.). В результате формируется неполноценный защитный гидролипидный слой, что также приводит к траскутанной потери воды и облегчению проникновения аллергенов и ирритантов.
3. Кожное сало. Функциональное значение кожного сала очень велико, выделяясь из секреторного отдела сальных желез, заполняя их выводные протоки и устья волосяных фолликул, секрет распределяется по бороздкам кожи и неравномерно покрывает практически всю ее поверхность слоем 7–10 мкм. За одну неделю у здорового человека выделяется 100–200 г секрета сальных желез, а при себорее 300 г и более. На поверхности кожи кожное сало смешивается с секретом потовых желез и эмульгируется. Таким образом, формируется тонкая водно-липидная эмульсионная пленка (sebum). Водно-липидная мантия подобно восковому налету предохраняет от избыточного солнечного излучения, переувлажнения, вредных воздействий внешней среды, инфекций, препятствует испарению воды и высвобождает глицерин, который связывает воду из атмосферного воздуха и удерживает у поверхности кожи [12, 13].
4. Кератин — конечный продукт жизнедеятельности эпидермиса, характеризуется стойкостью по отношению к механическим, физическим и химическим факторам. По мнению некоторых авторов, кератин, как все белки, является коллоидом — в воде набухает и связывает молекулы воды [14].
При нарушении одной или нескольких влагоудерживающих структур (дефицит компонентов, структурные изменения и т. д.) уровень воды в роговом слое падает. Происходит нарушение его структуры, что влечет за собой и нарушение барьерных свойств. Это означает, что роговой слой перестает быть непроницаемой преградой для воды и ее испарение усиливается. Через нарушенный барьер могут легче проникать микроорганизмы, химические факторы агрессии, которые дополнительно поддерживают раздражение и воспаление кожи. Сухость кожи — неизменный симптом различных кожных заболеваний, таких как атопический дерматит, псориаз, экзема и т. д. В последние годы стала преодолеваться необоснованная позиция среди практикующих врачей, касающаяся пренебрежения таким важным аспектом, как уход за кожей и восстановление ее барьерных функции у больных дерматозами. Поэтому во второй части статьи речь пойдет не только о вспомогательной терапии космецевтики во время обострения, но и об очень важном вопросе — закреплении ремиссии путем активного восстановления целостности кожи и ее нормальной функции с помощью лечебно-косметических средств.
Литература
- Ломакина Е. А. Роль барьерной функции кожи в патогенезе некоторых дерматозов // Современные проблемы дерматовенерологии, иммунологии и врачебной косметологии. 2009, № 2. С. 87–90.
- Калюжная Л. Д. Атопический дерматит и сухость кожи // Клиническая иммунология. Аллергология. Инфектология. 2009, № 1. С. 27–18.
- Ткаченко С., Эрнандес Е. Аквапорины в регуляции водного баланса кожи // Косметика и медицина. 2011, № 2. С. 26–33.
- Мядлец О. Д., Адаскевич В. П. Морфофункциональная дерматология. М.: Медлит, 2006. 752 с.
- Марголина А. А., Эрнандес Е. И., Зайкина О. Э. Новая косметология. М., 2002. 208 с.
- Кошевенко Ю. Н. Кожа человека. Т. 2. М.: Медицина, 2008. 754 с.
- Кошевенко Ю. Н. Кожа человека. Т. 1. М.: Медицина, 2006, 360 с.
- Современная наружная терапия дерматозов (с элементами физиотерапии) / Под. ред Н. Г. Короткого. Тверь: «Губернская медицина», 2001. 528 с.
- Cork M. J., Robinson D. A., Vasilopoulos Y. et al. New perspectives on epidermal barrier dysfunction in atopic dermatitis: gene-environment interactions // J Allergy Clin Immunol. 2006; 118 (1): 3–21.
- Dobrev H. Scientific Researches of the Union of Scientists // Series D. Medicine, Pharmacy and Stomatology. 2002; 1: 107–10. Plovdiv.
- Norlen L., Nicander I., Lundh Rozell B. et al. Inter- and intra-individual differences in human stratum corneum lipid content related to physical parameters of skin barrier function in vivo // J Invest Dermatol. 1999; 112 (1): 72–77.
- Roh M., Han M., Kim D., Chung K. Sebum Output as a Factor Contributing to the Size of Facial Pores // Br J Dermatol. 2006; 155 (5): 890–894.
- Аравийская Е. Р., Соколовский Е. В. Сухость кожи. Причины возникновения. Принципы коррекции // Журнал дерматовенерологии и косметологии. 2002, № 1. С. 23–25.
- Эрнандес Е. И. Увлажнение кожи. М.: ООО «Фирма Клавель», ООО «Школа косметических химиков», 2007. 32 с.
Ю. А. Галлямова, доктор медицинских наук, профессор
О. А. Баринова
РМАПО, Москва
Анатомия и физиология кожи
Кожа – наш самый большой орган, составляющий 15% от общей массы тела. Она выполняет множество функций, прежде всего защищает организм от воздействия внешних факторов физической, химической и биологической природы, от потери воды, участвует в терморегуляции. Последние научные данные подтверждают, что кожа не только обладает собственной иммунной системой, но и сама является периферическим иммунном органом.
Структура кожи
Кожа состоит из 3 слоев: эпидермиса, дермы и подкожной жировой клетчатки (ПЖК) (рис. 1). Эпидермис – самый тонкий из них, представляет собой многослойный ороговевающий эпителий. Дерма – средний слой кожи. Главным образом состоит из фибрилл структурного белка коллагена. ПЖК содержит жировые клетки – адипоциты. Толщина этих слоев может значительно варьировать в зависимости от анатомического места расположения.
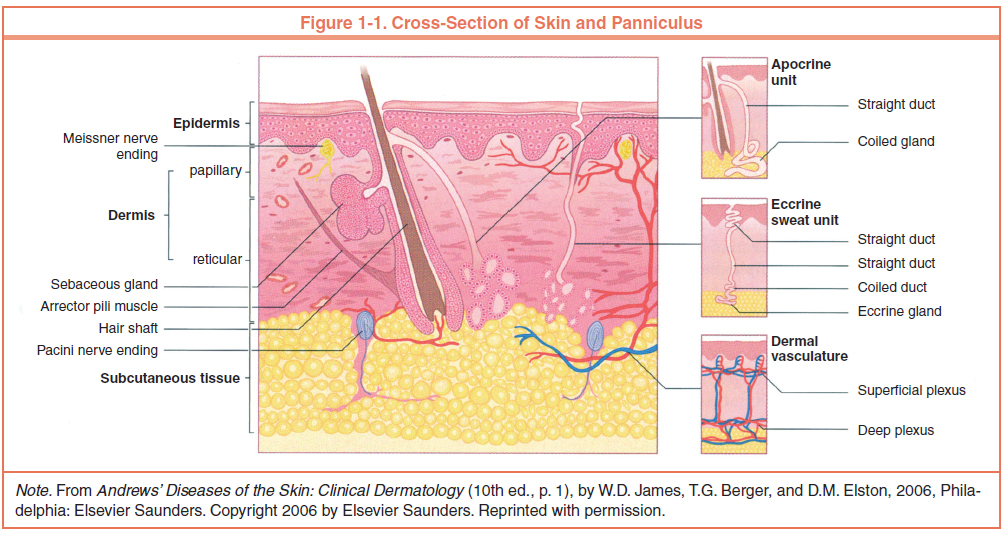
Рис.1. Структура кожи
Эпидермис
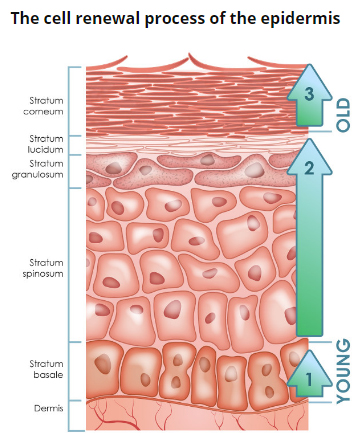
Рис. 2. Эпидермис
Эпидермис – постоянно слущивающийся эпителиальный слой кожи, в котором представлены в основном из 2 типа клеток – кератиноциты и дендритные клетки. В небольшом количестве в эпидермисе присутствуют меланоциты, клетки Лангерганса, клетки Меркеля, внутриэпидермальные Т-лимфоциты. Структурно эпидермис разделяется на 5 слоев: базальный, шиповатый, зернистый, блестящий и роговой , различающиеся положением и степенью дифференцировки кератиноцитов, основной клеточной популяции эпидермиса (рис. 2).
Кератинизация. По мере дифференцировки кератиноцитов и продвижения от базального слоя до рогового происходит их кератинизация (ороговевание) – процесс, начинающийся с фазы синтеза кератина кератиноцитами и заканчивающийся их клеточной деградацией. Кератин служит строительным блоком для промежуточных филаментов. Пучки из этих филаментов, достигая цитоплазматический мембраны, формируют десмосомы, необходимые для образования прочных контактов между соседними клетками. Далее, по мере процесса эпителиальной дифференцировки, клетки эпидермиса вступают в фазу деградации. Ядра и цитоплазматические органеллы разрушаются и исчезают, обмен веществ прекращается, и наступаетапоптозклетки, когда она полностью кератинизируется (превращается в роговую чешуйку).
Базальный слой эпидермиса состоит из одного ряда митотически активных кератиноцитов, которые делятся в среднем каждые 24 часа и дают начало новым клеткам новым клеткам вышележащих эпидермальных слоев. Они активируются только в особых случаях, например при возникновении раны. Далее новая клетка, кератиноцит, выталкивается в шиповатый слой, в котором она проводит до 2 недель, постепенно приближаясь к гранулярному слою. Движение клетки до рогового слоя занимает еще 14 дней. Таким образом, время жизни кератиноцита составляет около 28 дней.
Надо заметить, что не все клетки базального слоя делятся с такой скоростью, как кератиноциты. Эпидермальные стволовые клетки в нормальных условиях образуют долгоживущую популяцию с медленным циклом пролиферации.
Шиповатый слой эпидермиса состоит из 5-10 слоев кератиноцитов, различающихся формой, структурой и внутриклеточным содержимым, что определяется положением клетки. Так, ближе к базальному слою, клетки имеют полиэдрическую форму и круглое ядро, но по мере приближения клеток к гранулярному слою они становятся крупнее, приобретают более плоскую форму, в них появляются ламеллярные гранулы, в избытке содержащие различные гидролитические ферменты. Клетки интенсивно синтезируют кератиновые нити, которые, собираясь в промежуточные филаменты, остаются не связанными со стороны ядра, но участвуют в образовании множественных десмосом со стороны мембраны, формируя связи с соседними клетками. Присутствие большого количества десмосом придает этому слою колючий вид, за что он и получил название «шиповатый».
Зернистый слой эпидермиса составляют еще живые кератиноциты, отличающиеся своей уплощенной формой и большим количеством кератогиалиновых гранул. Последние отвечают за синтез и модификацию белков, участвующих в кератинизации. Гранулярный слой является самым кератогенным слоем эпидермиса. Кроме кератогиалиновых гранул кератиноциты этого слоя содержат в большом количестве лизосомальные гранулы. Их ферменты расщепляют клеточные органеллы в процессе перехода кератиноцита в фазу терминальной дифференцировки и последующего апоптоза. Толщина гранулярного слоя может варьировать, ее величина, пропорциональная толщине вышележащего рогового слоя, максимальна в коже ладоней и подошв стоп.
Блестящий слой эпидермиса (назван так за особый блеск при просмотре препаратов кожи на световом микроскопе) тонкий, состоит из плоских кератиноцитов, в которых полностью разрушены ядра и органеллы. Клетки наполнены элейдином – промежуточной формой кератина. Хорошо развит лишь на некоторых участках тела – на ладонях и подошвах.
Роговой слой эпидермиса представлен корнеоцитами (мертвыми, терминально-дифференцированными кератиноцитами) с высоким содержанием белка. Клетки окружены водонепроницаемым липидным матриксом, компоненты которого содержат соединения, необходимые для отшелушивания рогового слоя (рис. 3). Физические и биохимические свойства клеток в роговом слое различаются в зависимости от положения клетки внутри слоя, направляя процесс отшелушивания наружу. Например, клетки в средних слоях рогового слоя обладают более сильными водосвязывающими свойствами за счет высокой концентрации свободных аминокислот в их цитоплазме.
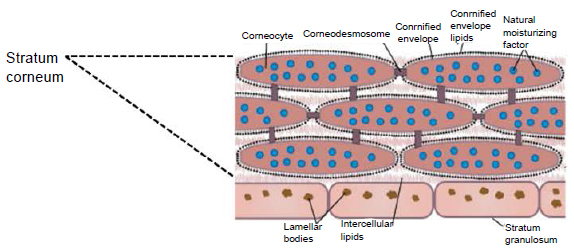
Рис. 3. Схематичное изображение рогового слоя с нижележащим зернистым слоем эпидермиса.
Регуляция пролиферации и дифференцировки кератиноцитов эпидермиса . Являясь непрерывно обновляющейся тканью, эпидермис содержит относительно постоянное число клеток и регулирует все взаимодействия и контакты между ними: адгезию между кератиноцитами, взаимодействие между кератиноцитами и мигрирующими клетками, адгезию с базальной мембраной и подлежащей дермой, процесс терминальной дифференцировки в корнеоциты. Основной механизм регуляции гомеостаза в эпидермисе поддерживается рядом сигнальных молекул – гормонами, факторами роста и цитокинами. Кроме этого, эпидермальный морфогенез и дифференцировка частично регулируются подлежащей дермой, которая играет критическую роль в поддержании постнатальной структуры и функции кожи.
Дерма
Дерма представляет собой сложноорганизованную рыхлую соединительную ткань, состоящую из отдельных волокон, клеток, сети сосудов и нервных окончаний, а также эпидермальных выростов, окружающих волосяные фолликулы и сальные железы. Клеточные элементы дермы представлены фибробластами, макрофагами и тучными клетками. Лимфоциты, лейкоциты и другие клетки способны мигрировать в дерму в ответ на различные стимулы.
Дерма, составляя основной объем кожи, выполняет преимущественно трофическую и опорную функции, обеспечивая коже такие механические свойства, как пластичность, эластичность и прочность, необходимые ей для защиты внутренних органов тела от механических повреждений. Также дерма удерживает воду, участвует в терморегуляции и содержит механорецепторы. И, наконец, ее взаимодействие с эпидермисом поддерживает нормальное функционирование этих слоев кожи.
В дерме нет такого направленного и структурированного процесса клеточной дифференцировки, как в эпидермисе, тем не менее в ней также прослеживается четкая структурная организация элементов в зависимости от глубины их залегания. И клетки, и внеклеточный матрикс дермы также подвергаются постоянному обновлению и ремоделированию.
Внеклеточный матрикс (ВКМ) дермы , или межклеточное вещество, в состав которого входят различные белки (главным образом коллаген, эластин), гликозаминогликаны, самым известным из которых является гиалуроновая кислота, и протеогликаны (фибронектин, ламинин, декорин, версикан, фибриллин). Все эти вещества секретируются фибробластами дермы. ВКМ представляет собой не беспорядочное скопление всех компонентов, а сложноорганизованную сеть, состав и архитектоника которой определяют такие биомеханические свойства кожи, как жесткость, растяжимость и упругость. К белкам ВКМ прикрепляются кератиноциты эпидермиса, которые тесно состыкованы друг с другом. Именно они и формируют плотный защитный слой кожи. Структура ВКМ также способна оказывать регулирующее влияние на погруженные в него клетки. Регуляция может быть как прямой, так и косвенной. В первом случае белки и гликозаминогликаны ВКМ непосредственно взаимодействуют с рецепторами клеток и инициируют в них специфические пути передачи сигнала. Косвенная регуляция осуществляется посредством действия цитокинов и ростовых факторов, удерживаемых в ячейках сети ВКМ и высвобождаемых в определенный момент для взаимодействия с рецепторами клеток. Структурная сеть ВКМ подвергается ремоделированию ферментами из семейства матриксных металлопротеиназ (ММР). В частности, ММР-1 и ММР-13 инициируют деградацию коллагенов I и III типов. Плотность сети ВКМ дермы неравномерна – в папиллярном слое она более рыхлая, в ретикулярном - значительно плотнее как за счет более близкого расположения волокон структурных белков, так и за счет увеличения диаметра этих волокон.
Коллаген – один из главных компонентов ВКМ дермы. Синтезируется фибробластами. Процесс его биосинтеза сложный и многоступенчатый, в результате которого фибробласт секретирует в экстрацеллюлярное пространство проколлаген, состоящий из трех полипептидных α-цепей, свернутых в одну тройную спираль. Затем мономеры проколлагена ферментивным путем собираются в протяженные фибриллярные структуры различного типа. Всего в коже не менее 15 типов коллагена, в дерме больше всего I, III и V типов этого белка: 88, 10 и 2% соответственно. Коллаген IV типа локализуется в зоне базальной мембраны, а коллаген VII типа, секретируемый кератиноцитами, играет роль адаптерного белка для закрепления фибрилл ВКМ на базальной мембране (рис. 4). Волокна структурных коллагенов I, III и V типов служат каркасом, к которому присоединяются другие белки ВКМ, в частности коллагены XII и XIV типов. Считается, что эти минорные коллагены, а также небольшие протеогликаны (декорин, фибромодулин и люмикан) регулируют формирование структурных коллагеновых волокон, их диаметр и плотность образуемой сети. Взаимодействие олигомерных и полимерных комплексов коллагена с другими белками, полисахаридами ВКМ, разнообразными факторами роста и цитокинами приводит к образованию особой сети, обладающей определенной биологической активностью, стабильностью и биофизическими характеристиками, важными для нормального функционирования кожи. В папиллярном слое дермы волокна коллагена располагаются рыхло и более свободно, тогда как ее ретикулярный слой содержит более крупные тяжи коллагеновых волокон.
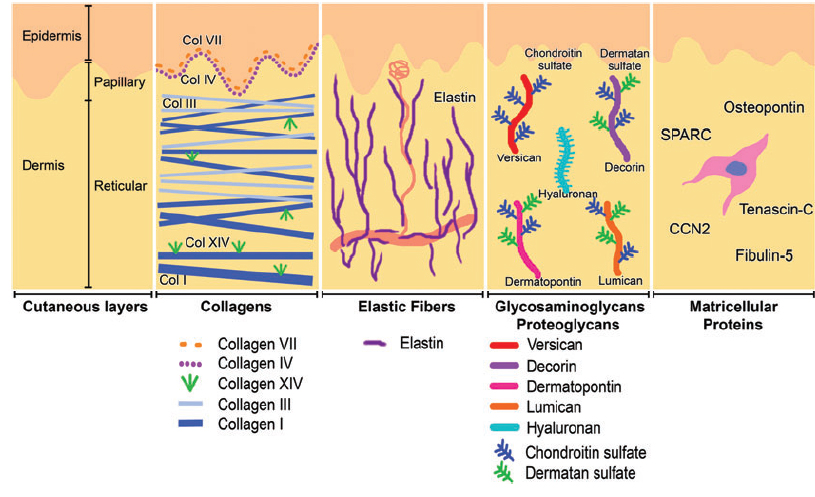
Рис. 4. Схематичное представление слоев кожи и распределения коллагенов разных типов.
Коллаген постоянно обновляется, деградируя под действием протеолитических ферментов коллагеназ и замещаясь вновь синтезированными волокнами. Этот белок составляет 70% сухого веса кожи. Именно коллагеновые волокна «держат удар» при механическом воздействии на нее.
Эластин формирует еще одну сеть волокон в дерме, наделяя кожу такими качествами, как упругость и эластичность. По сравнению с коллагеном эластиновые волокна менее жесткие, они скручиваются вокруг коллагеновых волокон. Именно с эластиновыми волокнами связываются такие белки, как фибулины и фибриллины, с которыми, в свою очередь, связывается латентный TGF-β-связывающий белок (LTBP). Диссоциация этого комплекса приводит к высвобождению и к активации TGF-β, самого мощного из всех факторов роста. Он контролирует экспрессию, отложение и распределение коллагенов и других матриксных белков кожи. Таким образом, интактная сеть из волокон эластина служит депо для TGF-β.
Гиалуроновая кислота (ГК) представляет собой линейный полисахарид, состоящий из повторяющихся димеров D-глюкуроновой кислоты и N-ацетилглюкозамина. Количество димеров в полимере варьирует, что приводит к образованию молекул ГК разного молекулярного веса и длины - 1х10 5 -10 7 Да (2-25 мкм), оказывающих, соответственно, различный биологический эффект.
ГК - высокогидрофильное вещество, влияющее на движение и распределение воды в матриксе дермы. Благодаря этому ее свойству наша кожа в норме и в молодости обладает высоким тургором и сопротивляемостью механическому давлению.
ГК с легкостью образует вторичные водородные связи и внутри одной молекулы, и между соседними молекулами. В первом случае они обеспечивают формирование относительно жестких спиральных структур. Во втором – происходит ассоциация с другими молекулами ГК и неспецифическое взаимодействие с клеточными мембранами, что приводит к образованию сети из полимеров полисахаридов с включенными в нее фибробластами. На длинную молекулу ГК, как на нить, «усаживаются» более короткие молекулы протеогликанов (версикана, люмикана, декорина и др.), формируя агрегаты огромных размеров. Протяженные во всех направлениях, они создают каркас, внося вклад в стабилизацию белковой сети ВКМ и фиксируя фибробласты в определенном окружении матрикса. В совокупности все эти свойства ГК наделяют матрикс определенными химическими характеристиками – вязкостью, плотностью «ячеек» и стабильностью. Однако сеть ВКМ является динамической структурой, зависящей от состояния организма. Например, в условиях воспаления агрегаты ГК с протеогликанами диссоциируют, а образование новых агрегатов между вновь синтезированными молекулами ГК (обновляющимися каждые 3 дня) и протеогликанами блокируется. Это приводит к изменению пространственной структуры матрикса: увеличивается размер его ячеек, меняется распределение всех волокон, структура становится более рыхлой, клетки меняют свою форму и функциональную активность. Все это сказывается на состоянии кожи, приводя к снижению ее тонуса.
Помимо регуляции водного баланса и стабилизации ВКМ, ГК выполняет важную регуляторную роль в поддержании эпидермального и дермального гомеостаза. ГК активно регулирует динамические процессы в эпидермисе, включая пролиферацию и дифференцировку кератиноцитов, окислительный стресс и воспалительный ответ, поддержание эпидермального барьера и заживление раны. В дерме ГК также регулирует активность фибробластов и синтез коллагена. Ремоделируя матрикс, ГК управляет функционированием клеток в матриксе, влияя на их доступность для различных факторов роста и изменяя их функциональную активности. От действия ГК зависит миграция клеток и иммунный ответ в ткани. Таким образом, изменения в распределении, организации, молекулярном весе и метаболизме ГК имеют значимые физиологические последствия.
Фибробласты представляют собой основной тип клеточных элементов дермы. Именно эти клетки отвечают за продукцию ГК, коллагена, эластина, фибронектина и многих других белков межклеточного матрикса, необходимых для формирования соединительной ткани. Фибробласты в различных слоях дермы различаются и морфологически, и функционально. От глубины их залегания в дерме зависит не только количество синтезируемого ими коллагена, но и соотношение типов этого коллагена, например I и III типов, а также синтез коллагеназы: фибробласты более глубоких слоев дермы производят меньшее ее количество. Вообще, фибробласты – очень пластичные клетки, способные менять свои функции и физиологический ответ и даже дифференцироваться в другой тип клеток в зависимости от полученного стимула. В роли последнего могут выступать и сигнальные молекулы, синтезированные соседними клетками, и перестройка окружающего ВКМ.
Подкожно-жировая клетчатка
Подкожно-жировая клетчатка , или гиподерма, - самый нижний слой кожи, располагается под дермой. Состоит из жировых долек, разделенных между собой соединительнотканными септами, содержащими коллаген и пронизанными крупными сосудами. Главными клетками жировых долек являются адипоциты, количество которых варьирует в различных областях тела. В настоящее время ПЖК рассматривают не только как энергетическое депо, но и как эндокринный орган, адипоциты которого участвуют в выработке ряда гормонов (лептина, адипонектина, резистина), цитокинов и медиаторов, оказывающих влияние на метаболизм, чувствительность к инсулину, функциональную активность репродуктивной и иммунной систем.
Внеклеточный матрикс (ВМК, англ. extracellular matrix, ECM) — внеклеточные структуры ткани (интерстициальный матрикс и базальные мембраны). Многокомпонентная субстанция, в которую погружены все клетки нашего организма. В последнее десятилетие интерес к внеклеточному матриксу значительно возрос. Это связано с установлением его роли в старении, клеточной дифференцировке, успешной терапии рака и лечении некоторых наследственных заболеваний.
Компоненты ВКМ синтезируют специализированные клетки. В соединительной ткани наиболее распространены фибробласты, но формировать ВКМ умеют и другие клетки: в хрящах, это хондроциты, а в костях — остеобласты. Компоненты матрикса могут синтезировать и клетки прилегающих органов: например, клетки эпителия сосудов производят компоненты рыхлой соединительной ткани.
Разрушители ВКМ. Порядок в ВКМ наводят белки — разрушители его компонентов. Наиболее важные из них, металлопротеиназы, «расчищают путь» клеткам, которые движутся в ВКМ, и уничтожают старые и «сломанные» компоненты матрикса.
Основное вещество ВКМ. Основу матрикса формируют гиалуроновая кислота и особые белки: гликопротеины и протеогликаны. В состав ВКМ входит и множество других белков со специфической функциональной нагрузкой.
В гликопротеинах доля углеводов не превышает 20%, углеводные цепи короткие, имеют нерегулярное строение и не содержат уроновых кислот. Это структурные белки, как коллаген и эластин. За счет самого распространенного структурного белка в организме — коллагена — ВКМ приобретает прочность, а за счет эластина — гибкость и эластичность.
Протеогликаны — сложные белки с высокой степенью гликозилирования, часто имеющие в своем составе уроновые кислоты. 90–95% массы такой молекулы составляют длинные углеводные цепи регулярного строения, а на белки приходится лишь 5–10%. Такое строение обуславливает высокую молекулярную массу протеогликанов. Протеогликаны запасают воду и полезные вещества. Выполняют функцию наполнителя (основного вещества). Благодаря полярной природе и сильному отрицательному заряду, они связывают катионы и основную часть воды. Играют роль межтканевых прослоек и смазочного материала в суставах.
Интегрины — трансмембранные клеточные рецепторы, которые взаимодействуют с матриксом и участвуют в межклеточной коммуникации. Благодаря этим контактам в клетках активируются сигнальные каскады, регулирующие экспрессию генов, отвечающие за пролиферацию и дифференцировку клеток, их выживание или апоптоз.
Межклеточный матрикс выполняет разнообразные функции:
- является основой соединительной ткани, её клетки образуют с веществами матрикса межклеточные контакты (гемидесмосомы, адгезивные контакты и др.), которые могут выполнять сигнальные функции и участвовать в локомоции клеток;
- обеспечивает механические контакты между клетками, образует механически прочные структуры (кости, хрящ, сухожилия и суставы) и транспорт химических веществ;
- составляет основу фильтрующих мембран (например, в почках);
- изолирует клетки и ткани друг от друга (например, обеспечивает скольжение в суставах и движение клеток);
- формирует пути миграции клеток, вдоль которых они могут перемещаться, например, при эмбриональном развитии.
Межклеточный матрикс имеет различные составные вариации и выполняет множество функций.

Клеточная мембана - это двойной слой липидов, большинство из которых – фосфолипиды. Интегрины, дистрогликаны и рецепторы домена дискоидина (DDR) – белки, пронизывающие мемрану клетки. Являются клеточными рецепторами, взаимодействуют с внешней средой и передают межклеточные сигналы.
Базальная мембрана разделяет клетку и соединительную ткань (матрикс). Значит, практически все клетки вступают в контакт с матриксом напрямую. Базальная мембрана сформирована ламинином (светлая пластинка) и коллагеном 4 типа (темная пластинка), они объединены белком нидогеном (энтактином), из этих компонентов создана пространственная структура, обеспечены механическая поддержка и защита клеток.
Фибронектин – белок клеточной адгезии, гликопротеин, также отвечающий за структуру ткани, может формировать мультимерные цепочки. Участвует в адгезии, то есть сцеплении клеток. Они помогают клеткам закрепляться на поверхностях и отвечают за их рост и перемещение в ВКМ.
Также в базальной мембране содержатся молекулы протеина перлекана, занимающегося поддержкой физиологического (эндотелиального) барьера между кровеносной системой и центральной нервной системой. Участвует в нейромышечном соединении, отвечая за доставку нервных импульсов к мышечным клеткам. Обеспечивает защиту нервной ткани от находящихся в крови микроорганизмов, токсинов, клеточных и гуморальных факторов иммунной системы, которые воспринимают нервную ткань как чужеродную.
Далее располагается межклеточный матрикс или соединительная ткань. Его составляющие:
Коллагены - белки, состоящие из остатков аминокислот (или пептидов). Молекула коллагена - это спираль из трёх закрученных аминокислотных (пептидных) цепочек. Эти цепочки объединяются в «жгуты» из цепочек — фибриллы, а из пучков фибрилл состоят как раз коллагеновые волокна. Фибриллярные белки, составляющие основу соединительной ткани организма (сухожилия, кости, хрящи, дерма и т.д.), их волокнами пронизан ВМК. Характерные свойства - прочность на разрыв, эластичность и гибкость. Эластичным белком с аналогичными свойствами является эластин.
Эластин формирует трехмерную сеть белковых волокон. Она обеспечивает механическую прочность ткани, обеспечивает контакты между клетками, формирует пути миграции клеток, вдоль которых они могут перемещаться, изолирует разные клетки и ткани друг от друга.
Аггрекан (протеогликановый хондроитинсульфат) – хрящевой специфичный протеогликановый ядерный белок. Связывает фибриллы коллагена II типа, удерживает и связывает воду, ГК и белки, формируя осмос, а значит, делает соединительную ткань устойчивой к большим нагрузкам. Области высокого содержания аггрекана и глюкозаминогликана способствуют осмосу, необходимому для нормального функционирования хрящевой ткани, создают «разбухание» ткани, которое препятствуют внешнему давлению на неё.
Гиалуроновая кислота (ГК) синтезируется встроенными в мембрану белками и затем «выдавливается» через нее в межклеточное пространство. ГК помогает интегринам проводить сигналы в клетку, регулирует клеточный ответ на эти сигналы и дает клеткам возможность закрепляться на различных поверхностях. Она участвует в регенерации ткани. Содержится во многих биологических жидкостях, в том числе синовиальной, отвечает за вязкость соединительной ткани. В связке с аггреканом формирует устойчивость к компрессии. Это основной компонент биологической смазки и суставного хряща, в котором присутствует в виде оболочки каждой клетки (хондроцита).
Коллаген 7 типа - связующий структурный компонент. Например, в коже это якорные фибриллы в связке дермы (собственно кожи) и эпидермиса. То есть он «скрепляет» и удерживает коллагеновые пучки IV-го типа (базальная мембрана, «держит в тонусе» эпидермис) и коллагеновые волокна I и III типов (основное пространство дермы).
Кожа состоит из эпидермиса и дермы, они отделены эпидермальной базальной мембраной. Слово «отделены» отражает только структурное разделение слоёв кожи. Но, правильнее пользоваться термином «дермо-эпидермальное соединение», которое отражает механическую и физиологическую взаимосвязь между двумя слоями, обеспечивая их жизнеспособность и целостность.
Дермо-эпидермальное соединение, или базальная мембрана, состоит из около 20 типов различных молекул и толщиной около 100 нанометров. Оно играет важную роль в поддержании структуры кожи, скрепляя эпидермис и дерму с помощью якорных комплексов. Контролирует пролиферацию и дифференцировку базальных клеток в эпидермисе, являясь проводником, как различных сигналов, так и всех питательных веществ в эпидермис из дермы.
Разрушение данного соединения приводит к появлению морщин, это происходит не только в коже пожилых людей. Дегенеративные изменения в структуре поверхности кожи также отмечаются на коже лица людей в возрасте от 20 до 30 лет. Считается, что такие повреждения и изменения являются признаками преждевременного старения кожи и приводят к образованию морщин и провисанию. Значит, ранний уход за базальной мембраной – необходимый и эффективный метод предотвращения старения кожи.
Основные причины возрастных изменений кожи:
- УФ-излучение.
- Обезвоживание.
- Стресс.
- Преждевременное старение:
- нарушение барьерных функций кожи;
- атака свободных радикалов;
- старение кожи, связанное с генетическими изменениями;
- -чувствительность, гиперчувствительность, мнимая чувствительность кожи.
Базальная мембрана - важнейшая структурная единица кожи, тонкий бесклеточный слой, отделяющий соединительную ткань от эпителия или эндотелия, представляет собой своеобразную границу между соединительной тканью и эпителием.
Состоит из двух пластинок: светлой (лат. lamina lucida) содержащей элементы, подобные протеинам, протеогликанам и антигену пузырчатки; тёмной (lamina densa), содержащей элементы, подобные коллагену, энтактину, гепарансульфату. Иногда к тёмной пластинке прилегает образование, называемое фиброретикулярной пластинкой (lamina fibroreticularis) , состоящей из коллагеновых фибрилл. Ретикулярная пластинка соединена с базальной пластинкой с помощью якорных фибрилл (коллаген типа VII) и микрофибрилл (фибриллин).
Светлая пластинка (lamina lucida/lamina rara) - толщина 20-30 нм, светлый мелкозернистый слой, прилежит к плазмалемме базальной поверхности эпителиоцитов. От полудесмосом эпителиоцитов вглубь этой пластинки, пересекая её, направляются тонкие якорные филаменты. Содержит протеины, протеогликаны и антиген пузырчатки.
Темная (плотная) пластинка (lamina densa) - толщина 50-60 нм, мелкозернистый или фибриллярный слой, расположен под светлой пластинкой, обращен в сторону соединительной ткани. В пластинку вплетаются якорные фибриллы, имеющие вид петель (образованы коллагеном VII типа), в который продеты коллагеновые фибриллы подлежащей соединительной ткани. Состав: коллаген IV, энтактин, гепарансульфат.
Ретикулярная (фиброретикулярная) пластинка (lamina reticularis) — состоит из коллагеновых фибрилл и микроокружения соединительной ткани, связанных с якорными фибриллами.
К функциям базальной мембраны относят:
- структурную;
- фильтрационную;
- миграционную (базальная мембрана формирует направление клеточных миграций);
- детерминирующую (базальная мембрана обеспечивает детерминацию полярности клеток);
- метаболизирующую;
- регенерирующую;
- морфогенетическую.
Состав:
- Коллаген IV типа - образует опорный каркас базальной мембраны. В случае, если коллаген IV типа находится в здоровом состоянии, то и мембрана функционирует правильно, держа оба слоя кожи вместе (соединительную ткань и эпидермис).
- Коллаген VII типа - представляет собой якорьки-скрепочки, плотно держащий и скрепляющий коллагеновые фибриллы (пучки) базальной мембраны с коллагеновыми фибриллами из дермы, т.е. коллаген VII-го типа «скрепляет» и удерживает коллагеновые пучки IV-го типа (базальная мембрана, которая «держит в тонусе» эпидермис), и коллагеновые волокна I и III типов (основное пространство дермы). В итоге, если всё функционирует и синтезируется вовремя, получаем плотно-сотканную кожную ткань, которую можно охарактеризовать, как «молодую»;
- гепарансульфат-протеогликан — компонент, участвующий в процессах клеточной адгезии, демонстрирующий ангигенные свойства;
- димеры — ключевой компонент фибрилл, обеспечивающий особую прочность мембраны;
- энтактин — компонент, связывающий коллаген в мембране с гликопротеинами;
- гликопротеины — адгезивный субстрат, посредством которого эпителиоциты фиксируются к мембране.
Функции:
- Именно от качества и целостности базальной мембраны зависит упругость и эластичность кожи.
- Базальная мембрана - это плотное бесклеточное образование, на котором располагаются клетки базального слоя эпидермиса, кератиноциты, имеющие с ней прочную связь.
- В ней протекают активные процессы внутриклеточного синтеза коллагена.
- Фильтр, не пропускающий крупные молекулы в глубокие слои кожи. Она выполняет механические, барьерные и обменные функции для эпидермиса, играет важную роль в контролировании клеточного поведения.
- Через нее осуществляется питание клеток базального слоя эпидермиса, а значит, улучшается его структура, укрепляется местный иммунитет.
- Через ее активизацию происходит улучшение обменных процессов в дерме, стимуляция выработки коллагена, что является основой для улучшения общего вида кожи.
- «Связной» между эпидермисом и дермой. Базальная мембрана связана с эпидермисом и дермой при помощи специализированных фибрилл-якорей, состоящих из молекул коллагена типа VII. Он играет роль «якоря» и осуществляет взаимосвязь структур дермы и эпидермиса.
К основному гликопротеину базальной мембраны относится ламинин-5. Находится в межклеточном материале, который связывает все внутренние органы в единый организм. Это, своего рода, основа, фундамент. Своими тремя короткими «веточками» она держится за другие такие же молекулы, создавая эту основу, а длинной присоединяется к клеткам, фактически прикрепляя органы к «основе». Ламинин жизненно необходим, потому что он практически «склеивает» все ткани, препятствует нашим телам разваливаться.
Ламинин 5 – это также пептид, передающий сигналы и активно участвующий в восстановлении кожи. При повреждении кожи он способствует ускоренной миграции и адгезии кератиноцитов для скорейшего закрытия дефекта, а значит, является незаменимым участником регенерации эпидермиса.
В современной эстетической медицине и косметологии есть много средств, действие которых направлено на восстановление эпидермиса, его ускоренное обновление и питание. Целые комплексы процедур нацелены на улучшение качеств дермы - усиление её упругости за счёт увеличения синтеза коллагена, эластина, гиалуроновой кислоты и др. Дермо-эпидермальное соединение (базальная мембрана), как отдельная важная структура кожи, должна награждаться особым вниманием специалистов при выборе процедур для омоложения кожи.
В представленной статье обобщены современные данные о строении и функции эпидермального барьера, рассмотрены вопросы его роли в развитии атопического дерматита.
This article summarizes current data on the structure and function of the epidermal barrier, we also considered its role in development of atopic dermatitis.
С тех пор как наземная жизнь стала угрожать млекопитающим обезвоживанием, все более становятся актуальными исследования структурных, клеточных, биохимических и регуляторных механизмов организма, участвующих в поддержании гомеостаза проницаемости эпидермального барьера [1].
Кожа обеспечивает жизненно важную функцию, защищая позвоночных от повседневных и экстремальных влияний факторов внешней среды, таких как воздействие антигенов, ультрафиолетового света, моющих средств, микроорганизмов, токсинов, наночастиц и иных повреждений [2]. Таким образом, эпидермальный барьер, являясь физическим барьером [3], обеспечивает защиту организма на границе двух сред, ограничивая как потерю воды из организма, так и попадание патогенных микроорганизмов в организм [4].
Важнейшая составная часть эпидермального барьера, от которой зависит проникновение раздражителей и аллергенов, находится в нижней части рогового слоя [5]. Несмотря на минимальные размеры (толщина около 7–35 мкм), он играет определяющую роль в формировании защитного барьера и способствует предотвращению чрезкожного проникновения патогенов. Кроме роли физического барьера, роговой слой участвует в терморегуляции, газообмене, гидратации, обеспечении врожденного иммунитета, а создавая определенную pH поверхности кожи (слегка кислую), обеспечивает дополнительную защиту от болезнетворных микроорганизмов [6].
Во время терминальной дифференцировки кератиноциты зернистого слоя «заменяют» свою клеточную мембрану на нерастворимый белковый слой — роговую оболочку [7]. Она придает прочность корнеоцитам и является «платформой» для прикрепления липидов. Роговая оболочка в основном сформирована структурными белками, такими как лорикрин, инволюкрин, филаггрин (ФЛГ) и малыми пролин-богатыми белками. Особенно важен филаггрин, объединяющий кератиновые волокна клеток, «скручивая» кератиноциты в уплощенные диски с большой площадью поверхности [8].
Вокруг корнеоцитов расположены холестерин, фосфолипиды и церамиды, создавая многослойную мембрану из липидных пластинок [3, 9]. Последние играют важную роль в регуляции водного гомеостаза кожи, помогают сохранять воду в организме, а также предотвращают проникновение патогенных микроорганизмов и аллергенов [10].
На границе между роговым и зернистым слоями происходит секреция ламеллярных телец, содержащих предшественники липидов, а также необходимые ферменты для переработки этих прекурсоров в зрелые составляющие липидной матрицы [11]. Перед секрецией происходит сборка ламеллярных телец в пластинчатом комплексе, посредством дифференцированной их упаковки с различным содержимым [12]. Кроме вышеупомянутых метаболитов липидов, липидные тельца доставляют в межклеточное пространство рогового слоя протеолитически активные ферменты, такие как сериновые протеазы семейства калликреина, а также их ингибиторов, в том числе и лимфоэпителиальный ингибитор сериновых протеаз типа Kazal-5 (LEKTI) [13].
Единство между корнеоцитами рогового слоя зависит прежде всего от корнеодесмосом. Подобно десмосомам, они поддерживают целостность тканей посредством молекул межклеточной адгезии с помощью кальций-зависимых взаимодействий между двумя представителями семейства кадгерина, внеклеточных трансмембранных гликопротеинов — десмоглеина и десмоколлина. Внутри корнеоцитов десмоглеин и десмоколлин соединяют кератиновые филламенты корнеодесмосом посредством «соединительной бляшки», состоящей из плакоглобина, десмоплакина и плакофиллина. Десмоглеин и десмоколлин перемещаются от оболочки корнеоцитов в липидную пластину между корнеоцитами и связывают воедино белки соседних клеток. Кроме того, в процессе кератинизации эпителия экспрессируется специфический белок с молекулярной массой 52 кДа — корнеодесмосин [8, 14]. После секреции в экстрацеллюлярное пространство корнеодесмосин перемещается в область между зернистым и роговым слоями и проникает в десмосому. Появление корнеодесмосина знаменует переход от десмосом к корнеодесмосомам [3, 12].
Во время «нормальной» десквамации рогового слоя самые поверхностные корнеоциты «уходят» с поверхности кожи, замещаясь кератиноцитами, подвергшимися терминальной дифференцировке [14, 15]. Таким образом, существует «тонкий» баланс между базальной клеточной пролиферацией и десквамацией рогового слоя. Это обеспечивает постоянное обновление клеток эпидермиса, а также способствует поддержанию одинаковой толщины эпителия [14, 16]. Caubet и соавт. [15] описали несколько деградационных протеаз, которые разрушают внеклеточные корнеодесмосомальные адгезивные белки, связывающие корнеоциты вместе.
В роговом слое среди протеаз, вовлеченных в процесс десквамации, присутствуют хемотрипсиновые (SCCE) и трипсиновые ферменты (SCTE), которые экспрессируются кератиноцитами зернистого слоя и находятся во внеклеточном пространстве рогового слоя [14]. Данные протеазы гидролизуют корнеодесмосин и десмоколлин [15]. Их активность контролируется семейством специфических ингибиторов, в которое входит LEKTI [17]. При этом скорость десквамации коррелирует с количеством ингибиторов протеаз, поступивших из ламеллярных телец [3].
На барьерную функцию эпидермиса влияют структура, дифференцировка, функциональная активность кератиноцитов, качественный и количественный состав липидов рогового слоя эпидермиса, наличие веществ, относящихся к естественному увлажняющему фактору, а также рН поверхности кожи [18]. При изменении каких-либо из перечисленных показателей барьерная функция эпидермиса нарушается, что приводит к усиленному испарению воды через роговой слой. У человека эпидермальный барьер восстанавливается на 60% через 12 часов, а полное восстановление занимает 72 часа [19]. Полагают, что быстрое восстановление эпидермального барьера происходит за счет массового выброса из клеток готовых ламеллярных телец. В медленную фазу восстановления усиливается синтез жирных кислот, холестерина, церамидов, а также образование новых ламеллярных гранул. Нарушение восстановления кожного барьера приводит к обезвоживанию эпидермиса и развитию выраженной сухости кожи, что и наблюдается при развитии атопического дерматита (АД) [20].
Повреждение эпидермального барьера способствует проникновению через кожу микробов, аллергенов и различных поллютантов (токсины, раздражающие и загрязняющие вещества). Проникновение агентов с антигенными свойствами увеличивает риск сенсибилизации, поскольку взаимодействие аллергенов и аллерген-представляющих клеток кожи инициирует воспаление [14]. Степень дисфункции барьера коррелирует со степенью воспаления и тяжестью АД [11]. Впервые Y. Werner и M. Lindberg (1985 г.) привели доказательства нарушений барьерной функции кожи, измерив трансэпидермальную потерю воды (ТЭПВ) у лиц с АД. Авторам удалось показать, что у больных АД нарушения эпидермального барьера отмечаются не только в очагах активного воспаления, но и на клинически неизмененной коже [10, 21].
Изменения в свойствах эпидермального барьера, которые наблюдаются при АД, включают увеличение ТЭПВ, изменения рН поверхности кожи, повышение ее проницаемости, увеличение бактериальной колонизации, изменение в экспрессии антимикробных пептидов, а также нарушение его целостности. При аномалии эпидермального барьера аллергены, раздражающие и другие вещества могут проникать через кожу, способствуя обострению АД [6].
В настоящее время нарушение функции кожного барьера является одним из ведущих этиопатогенетических факторов АД. По данным разных авторов у 30–66% больных АД протекает без изменения уровня общего IgE в сыворотке крови и специфичных IgE к аллергенам. Полагают, что именно у этих пациентов нарушение функции кожного барьера играет ведущую роль в развитии и прогрессировании заболевания [22].
В последние годы определен ряд генетических мутаций, приводящих к изменениям в роговом слое эпидермиса. Так, у больных АД определяются мутации гена профилаггрина [23], способствующие нарушению синтеза белка ФЛГ — одного из компонентов цитокератинового скелета, обеспечивающего ороговение кератиноцитов. В результате изменения терминальной дифференцировки кератиноцитов развивается их функциональная несостоятельность [24]. Кроме того, снижение экспрессии гена ФЛГ уменьшает уровень «натурального увлажняющего фактора» [25]. Дефицит ФЛГ и/или продуктов его деградации способствует сухости кожи, что коррелирует с клинической картиной нарушения эпидермального барьера при АД [26]. Более того, при уменьшении ФЛГ усиливается «раздражающий» эффект аллергенов на дендритные клетки, понижая при этом порог их чувствительности к воспалению, что демонстрирует важность этого белка в предотвращении пенетрации аллергенов [27].
Кроме того, у больных АД выявлены полиморфные варианты гена SPINK5, приводящие к дизрегуляции синтеза ингибитора сериновой протеазы LEKTI, который в норме блокирует избыточную продукцию SCCE [28]. Активность деградационных протеаз также значимо связана с дефектом кожного барьера при АД [23]. Разбалансировка функционирования этих ферментов ведет к усиленному отшелушиванию клеток и истончению рогового слоя, увеличивая проницаемость эпидермального барьера [14]. Установлено, что у больных АД изменена липидная мантия рогового слоя, предупреждающая ТЭПВ. Отмечается низкий уровень церамидов первого и третьего типов в сочетании с повышением уровня свободного холестерола [29]. Повышение уровня рН на поверхности кожи приводит к снижению синтеза липидов и активизации ферментов (в частности, SCCE), повреждающих корнеодесмосомы. Уменьшение количества веществ, относящихся к «натуральному увлажняющему фактору», также способствует нарушению функционирования эпидермального барьера [30].
Таким образом, у больных АД отмечается комплекс нарушений, приводящих к дисфункции кожного барьера, усилению ТЭПВ и обезвоживанию кожных покровов [14]. Из вышеуказанного следует, что пациенты с АД могут иметь дефект кожного барьера, в основе которого лежат как генетические, так и приобретенные механизмы. Этот дефект способны спровоцировать внешние факторы, такие как царапины, использование моющих средств, микробная колонизация, а также воздействие протеаз различных аллергенов (в частности, протеаз клещей домашней пыли) [10]. Распознание конкретных механизмов, участвующих в патогенезе воспалительных заболеваний кожи, в частности, АД, безусловно, будет способствовать поиску и разработке новых, более эффективных методов воздействия на патологический процесс с минимальным количеством системных эффектов [8].
Литература
- Elias P. M. The skin barrier as an innate immune element // Semin. Immunopathol. 2007. Vol. 29 (1); р. 3–14.
- De Benedetto A., Kubo А., Beck L. A. Skin barrier disruption — a requirement for allergen sensitization? // J. Invest. Dermatol. 2012. Vol. 132 (3); р. 949–63.
- Danby S. G., Cork M. J. New understanding of atopic dermatitis: the role of epidermal barrier dysfunction and subclinical inflammation. // J. Clin. Dermatol. 2010. Vol. 1 (2); р. 33–46.
- Котлуков В. К., Кузьменко Л. Г., Антипова Н. В. Наружные дерматологические средства серии «Бепантен» в практике педиатра // Медицинский совет. 2012. Т. 5; с. 81–85.
- Madison K. С. Barrier function of the skin: «la raison d’etre» of the epidermis // J. Invest. Dermatol. 2003. Vol. 121; р. 231–241.
- Lorena S. T., Morello III A. P., Mack Correa M. C. et al. The infant skin barrier: can we preserve, protect, and enhance the barrier? // Dermatol. Res. Pract. 2012. Doi: 10.1155/2012/198789.
- Candi E., Schmidt R., Melino G. The cornified envelope: a model of cell death in the skin // Nat. Rev. Mol. Cell Biol. 2005. Vol. 6; р. 328–340.
- Michael J. C., Simon G. D., Yiannis V. et al. Epidermal barrier dysfunction in atopic dermatitis // J. Invest. Dermatol. 2009. Vol. 129; р. 1892–908.
- Ревякина В. А. Современные тенденции в комплексной наружной терапии атопического дерматита у детей // Лечащий Врач. 2014. № 6, с. 38.
- De Benedetto А., Agnihothri R., McGirt L. Y. et al. Atopic dermatitis: a disease caused by innate immune defects? // J. Invest. Dermatol. 2009. Vol. 129; р. 14–30.
- Roelandt T., Thys B., Heughebaert C. et al. LEKTI-1 in sickness and in health // Int. J. Cosmetic Sci. 2009. Vol. 31 (4); р. 247–254.
- Ishida-Yamamoto A., Simon M., Kishibe M. et al. Epidermal lamellar granules transport different cargoes as distinct aggregates // J. Invest. Dermatol. 2004. Vol. 122; р. 1137–1144.
- Ishida-Yamamoto A., Deraison C., Bonnart C. et al. LEKTI is localized in lamellar granules, separated from KLK5 and KLK7, and is secreted in the extracellular spaces of the superficial stratum granulosum // J. Invest. Dermatol. 2005. Vol. 124; р. 360–366.
- Cork M. J., Robinson D. A., Vasilopoulos Y. et al. New perspectives on epidermal barrier dysfunction in atopic dermatitis: gene-environment interactions // J. Allergy Clin. Immunol. 2006. Vol. 118; р. 3–21.
- Caubet C., Jonca N., Brattsand M. et al. Degradation of corneodesmosome proteins by two serine proteases of the kallikrein family, SCTE/KLK5/hK5 and SCCE/KLK7/hK7 // J. Invest. Dermatol. 2004. Vol. 122; Р. 1235–1244.
- Goldstein A. M., Abramovits W. Ceramides and the stratum corneum: structure, function, and new methods to promote repair // Int. J. Dermatol. 2003. Vol. 42; р. 256–59.
- Chavanas S., Bodemer C., Rochat A. Mutations in SPINK5, encoding a serine protease inhibitor, cause Netherton syndrome // Nat. Genet. 2000. Vol. 25; р. 141–142.
- Мурашкин Н. Н. Нарушения дермального барьера как фактор хронизации дерматозов в детском возрасте, подходы к терапии: теоретические и практические инновации // Лечащий Врач. 2014. № 6, с. 19.
- Эрнандес Е., Марголина А., Петрухина А. Липидный барьер кожи и косметические средства // Косметика и медицина. 2005. Т. 6; с. 35–38.
- Тамразова О. Б. Синдром сухой кожи // Дерматология. Приложение к журналу Consilium Medicum. 2014. Т. 3; с. 25–31.
- Хлебникова А. Н. Увлажняющие средства в терапии хронических дерматозов // Клиническая дерматология и венерология. 2010. Т. 4; с. 32–39.
- Palmer C. N., Irvine A. D., Terron-Kwiatkowski A. et al. Common loss-of-function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis // Nat. Genet. 2006. Vol. 38 (4); р. 441–446.
- Молочков А. В., Тамразова О. Б. Ксероз кожи — основной патогенетический фактор развития атопического дерматита // Дерматология. Приложение к журналу Consilium Medicum. 2014. Т. 4; с. 48–54.
- Vasilopoulos Y., Cork M. J., Murphy R. et al. Genetic association between an AACC insertion in the 3’UTR of the stratum corneum chymotryptic enzyme gene and atopic dermatitis // J. Invest. Dermatol. 2004. Vol. 123; р. 62–66.
- Smith F. J., Irvine A. D., Terron-Kwiatkowski A. et al. Loss-of-function mutations in the gene encoding filaggrin cause ichthyosis vulgaris // Nat. Genet. 2006. Vol. 38 (3); р. 337–342.
- Nemoto-Hasebe I., Akiyama M., Nomura T. et al. Clinical severity correlates with impaired barrier in? laggrin-related eczema // J. Invest. Dermatol. 2009. Vol. 129 (3); р. 682–689.
- Fallon P. G., Sasaki T., Sandilands A. et al. A homozygous frameshift mutation in the mouse Flg gene facilitates enhanced percutaneous allergen priming // Nat. Genet. 2009. Vol. 41 (5); р. 602–608.
- Kato A., Fukai K., Oiso N. et al. Association of SPINK5 gene polymorphisms with atopic dermatitis in the Japanese population // Br. J. Dermatol. 2003. Vol. 148; р. 665–669.
- Пампура А. Н., Чусляева А. А. Современные подходы к терапии атопического дерматита у детей // Медицинский вестник Башкортостана. 2014. Т. 9 (1); с. 93–98.
- Eberlein-Konig B., Schafer T., Huss-Marp J. et al. Skin surface pH, stratum corneum hydration, transepidermal water loss and skin roughness related to atopic eczema and skin dryness in a population of primary school children // Acta Dermatol. Venereol. 2000. Vol. 80; р. 188–191.
С. В. Левашева 1 , кандидат медицинских наук
Э. И. Эткина, доктор медицинских наук, профессор
Л. Л. Гурьева, кандидат медицинских наук
Л. И. Бабенкова, кандидат медицинских наук
Л. Я. Данилова
С. Э. ЯкутаЧитайте также: