
Адгезия на эпителии что это
Обновлено: 26.04.2024
Адгезия. Поглощение микробов. Схема фагоцитоза.
Адгезия [от лат. adhaesio, приклеиваться]. Одно из условий успешного поглощения возбудителя — эффективная адгезия к микробу. Жгутики позволяют микробам быстро перемещаться в жидкой фазе, а фагоциты не умеют «плавать», но хорошо бегают, то есть свои поглотительные свойства они способны реализовывать только на какой-либо плотной поверхности (например, на эпителии). Опсонины [от греч. opson, лакомство], такие как AT, C3b, фибро-нектин, сурфактант, обволакивают микроорганизмы и существенно ограничивают их подвижность. Опсонины делают поглощение более эффективным, что связано со стабильностью взаимодействий опсонинов с соответствующими рецепторами (к Fc-фрагментам ЛТ, компонентам комплемента, фибронектину и др.) на мембране фагоцита (рис. 10~3). Отсутствие этих рецепторов приводит к резкому снижению функциональной активности фагоцитов (например, врождённый дефицит С3b-рецепторов сопровождается высокой частотой бактериальных инфекций и даже выделен в отдельную нозологическую форму — недостаточность адгезии лейкоцитов).
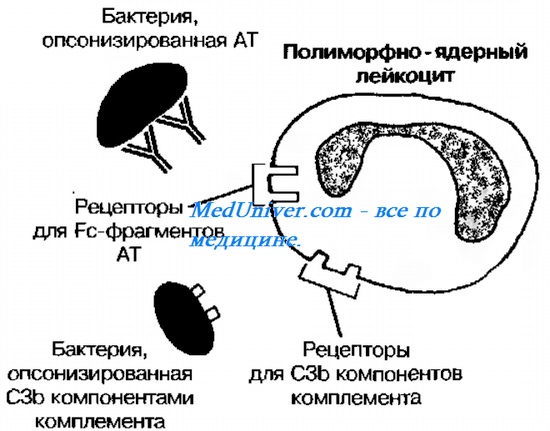
Рис. 10-3. Фагоцитоз. Схема фагоцитоза. Схема участия опсонинов в фагоцитарных реакциях. Бактерии опсонизируются молекулами AT и С3b-компонента комплемента, рецепторы к которым экспрессируются на поверхности фагоцитов. Взаимодействие соответствующих рецепторов с лигандами облегчает поглощение бактерии в ходе фагоцитоза.
Поглощение микробов
Поглощение микробов идентично таковому у амёб; в результате образуется фагосома с заключённым внутри объектом фагоцитоза (рис. 10-4, А). К фагосоме устремляются лизосомы и выстраиваются по её периметру. Затем мембраны фагосомы и лизосом сливаются (фагосомо-лизосомальное слияние), и ферменты лизосом изливаются в образовавшуюся фаголизосому. Поглощению способствует взаимодействие поверхностных рецепторов фагоцитов с Аг или фрагментами опсонинов, сорбированных на поверхности бактерии. Эта реакция напоминает действие замка-молнии (так называемый зипперный механизм поглощения [от англ. zip, замок-молния]).
Фагоцитированные микроорганизмы подвергаются атаке комплекса различных микробицидных факторов, разделяемых на кислородзависимые и кислороднезависимые.
- Вернуться в оглавление раздела "Микробиология."
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
ВЛИЯНИЕ ПРЕБИОТИКОВ НА ПРОЦЕССЫ АДГЕЗИИ И КОГЕЗИИ БИФИДОБАКТЕРИЙ

Одним из актуальных направлений современной микробиологии является изучение адгезивного процесса различных микроорганизмов. Известно, что способность микроорганизмов приживаться в ЖКТ, создавая защитный барьер, обусловлена их адгезивными свойствами. Адгезия - это межклеточное взаимодействие, выражающееся в прочном прикреплении клеток к субстрату. Следует отметить, что от адгезивных свойств во многом зависят стабильность и защитные свойства микрофлоры макроорганизма.
Адгезия к слизи, гликопротеинам и эпителиальным клеткам, а также колонизация в желудочно-кишечном тракте человека является основополагающими характеристиками микроорганизмов, обладающих пробиотическими свойствами.
Злаковые культуры и жмых ядра кедрового ореха содержат высокое количество растворимых и нерастворимых пищевых волокон и могут адсорбировать и колонизировать бифидобактерии в процессе культивирования. В связи с этим в дальнейших исследованиях изучали адгезивные свойства бифидобактерий при их культивировании на питательной среде с внесением пребиотиков. Адгезию бифидобактерий оценивали по среднему показателю адгезии (СПА), коэффициенту участия эритроцитов (КУЭ), индексу адгезивности микроорганизмов (ИАМ). Микроорганизмы считаются высокоадгезивными - при ИАМ более 4,0; среднеадгезивными – при ИАМ от 2,51 - 4,0; малоадгезивными – при ИАМ от 1,76 – 2,54. Результаты исследований представлены в таблице 1.
Таблица 8 - Влияние пребиотиков на адгезивные свойства бифидобактерий В. longum ДК-100 и B. bifidum 83
Результаты исследований, представленные в таблице 8, свидетельствуют о том, что исследуемые штаммы бифидобактерий в присутствии пребиотиков обладают более высокими адгезивными свойствами.
Необходимо отметить, что бактериальная поверхность заряжена отрицательно, у грамположительных бактерий это обусловлено присутствием тейхоевых кислот. В результате физико-химических взаимодействий, как электростатических, так и прочих, микробная клетка притягивается к поверхности (субстрату), и таким образом возникает начальный контакт. «Субстрат» покрыт слоем высокомолекулярных полисахаридов, пребиотиков. Сближение микробной клетки с субстратом вызывает изменение ее формы и перераспределение заряженных и незаряженных групп на контактирующей клеточной поверхности. В результате чего зона контакта увеличивается, что способствует взаимодействию с субстратом. Обычно в процессе прикрепления к поверхности синтезируются адгезины, о чем свидетельствует полученные нами результаты (табл. 8).
Прикрепление также может быть инициировано гидрофобными взаимодействиями. Поскольку все молекулы, и полярные и неполярные, в результате физико-химического взаимодействия накапливаются у границы с поверхностью, прикрепленные бактерии лучше снабжаются необходимыми веществами, чем свободные клетки в жидкой среде. На прикрепленные бактерии в меньшей степени воздействуют токсичные соединения, в том числе кислород, так как они диффундируют только с одной стороны и нейтрализуются всем микробным сообществом, а не одиночными клетками. Это явление особенно важно для анаэробных микроорганизмов, к которым относятся бифидобактерии.
Прикрепленные бактерии гораздо легче, чем свободноплавающие бактерии, образуют кооперативные структуры друг с другом или с другими бактериями. В прикрепленных бактериальных сообществах имеются также большие возможности для обмена плазмидами.
Вероятно, высокомолекулярные полисахариды, содержащиеся в исследуемых растительных добавках и синтез адгезинов способствуют прикреплению бифидобактерий к пищевым волокнам.
К механизмам, гарантирующим стабильность микробного консорциума, кроме адгезии относится также когезия (агрегация клеток). В литературных источниках недостаточно сведений о межклеточных контактах микроорганизмов, отражающих закономерности развития микробных популяций как саморегулирующих многоклеточных систем.
Известно, что популяции бифидобактерий демонстрируют высокоорганизованное, мицелиоподобное развитие на искусственных питательных среда. Топография взаиморасположения палочковидных, коккоидных и мультисиптированных форм бифидобактерий в мицелии определяется вектором репродуктивных потенций, а также обособлением клеток в процессе формирования перегородок и разделения дочерних особей. В литературе имеются единичные сведения, освещающие агрегацию клеток бифидобактерий и образованию в дальнейшем сложных многоклеточных систем.
Следующий этап исследований посвящен изучению влияния пребиотиков на когезию бифидобактерий. Результаты исследований представлены на рисунках 4-5.
Строение и свойства адгезивных контактов клеток
• Адгезивные контакты представляют собой семейство близких поверхностных доменов, которые связывают соседние клетки друг с другом
• Адгезивные контакты содержат трансмембранные кадгериновые рецепторы
• Наиболее полно изученный адгезионный контакт, называемый опоясывающим зональным контактом, или адгезивным пояском (zonula adherens), расположен в контактном комплексе, который образуется в некоторых тканях между соседними эпителиальными клетками
• В опоясывающем зональном контакте адаптерные белки, которые называются катенинами, связывают кадгерины с актиновыми филаментами
Адгезивные контакты представляют собой компоненты контактного комплекса, благодаря которому скрепляются между собой клетки эпителия и эндотелия. В электронном микроскопе адгезивные контакты выглядят в виде темных утолщенных полос, расположенных в примыкающих областях плазматической мембраны соседних клеток, связанных стержнеобразными структурами, простирающимися в межклеточное пространство. На представлен наиболее полно исследованный адгезивный контакт, называемый опоясывающий зональный контакт, или адгезивный поясок (zonula adherens) В комплексе, формирующемся между некоторыми эпителиальными клетками, он располагается непосредственно под плотным контактом.
(Исследователи, впервые обнаружившие этот контакт, назвали его «опоясывающей десмосомой» из-за внешнего сходства с большой десмосомой. Сейчас мы знаем, что этот контакт отличается от десмосомы, и поэтому старое название больше не используется.) Другие примеры адгезивных контактов включают контакт в синапсе между нейронами ЦНС, контакт во вставочных дисках между клетками миокарда, и контакт между слоями миелиновой оболочки, окружающей периферические нервы.
Несмотря на свое местоположение, адгезивные контакты имеют два общих свойства. Во-первых, они содержат трансмембранные белки-рецепторы, которые называются кадгерины. Эти белки связываются с одноименными белками, расположенными на соседних клетках. Связывание рецепторов, расположенных на одной клетке, с такими же рецепторами другой называется гомофильным связыванием.
Предполагается, что этот тип связывания играет важную роль в обеспечении клеточной организации ткани, способствуя возникновению специфического взаимодействия клеток между собой. Это можно проиллюстрировать, экспрессируя гены двух различных кадгериновых рецепторов в двух разных популяциях одних и тех же клеток, поместив эти популяции в одну чашку Петри. Через несколько часов клетки самоорганизуются в группы, экспрессирующие тот или иной рецептор.
Если к культуре добавить антитела, блокирующие сайты гомофильного связывания кадгеринов, то группы клеток не образуются.
Димеры кадгериновых рецепторов, участвующие в адгезивных контактах, содержат пять внеклеточных доменов, которые полностью обеспечивают гомофильное связывание. Как показано на рисунке ниже, возможны три отдельных перекрывающихся положения этих доменов. Наиболее прочное связывание происходит, когда рецепторы перекрываются антипараллельно и полностью, в то время как их частичное перекрывание обеспечивает более слабые взаимодействия.
При изменении числа кадгериновых рецепторов на поверхности клеток может меняться прочность их связывания с соседними клетками; этот процесс называется модуляцией связывания. Мы не располагаем данными в пользу того, что кадгерины претерпевают конформационные изменения, приводящие к изменению сродства и контролирующие прочность связывания адгезивных контактов. Такие конформационные изменения известны под названием модуляции сродства и характерны для интегриновых рецепторов.
Второе свойство адгезивных контактов связано с тем, что они оказываются достаточно сильными для того, чтобы изменять форму ткани и/или противостоять силам сдвига. Например, как схематически показано на рисунке ниже, в связывании цитоплазматических хвостов кадгериновых рецепторов с пучками актина в опоясывающем зональном контакте участвуют якорные белки, известные под названием катенины. В свою очередь, актиновые филаменты этих пучков присоединены к миозиновым белкам, которые обеспечивают им возможность скользить по отношению друг к другу.
Считается, что это приводит к сокращениям, меняющим форму апикального полюса эпителиальных клеток. Это может быть важно, например, при развитии нервной трубки, когда за счет инвагинации эпителиальных клеток закрывается ее желобок.
Наряду с адгезивными функциями, обусловленными присутствием кадгерина, адгезивные контакты также участвуют в специфических процессах неизвестной природы. На основании генетических экспериментов на плодовой мушке предполагается, что в формировании морфологически четких опоясывающих зональных контактов участвуют белки, отличные от кадгеринов и катенинов. Эти дополнительные белки могут также регулировать сборку цитоскелета на сайтах, расположенных на значительных расстояниях от самих адгезивных контактов. Например, они могут участвовать в формировании полярности эпителиальных клеток, и, таким образом, косвенно влиять на сборку других клеточных контактов, включая плотные. В настоящее время исследователи пытаются выяснить, каким образом это происходит.
Опоясывающий зональный контакт является частью контактного комплекса.
С цитоплазматической стороны плазматической мембраны к контакту примыкают актиновые пучки.
Кадгерины образуют стержневые структуры между клетками и связываются с актиновым цитоскелетом через якорные белки, такие как катенины. Каждый тип адгезивных контактов прочно удерживает соседние клетки вместе Клетки, экспрессирующие идентичные кадгериновые рецепторы, селективно связываются между собой.
Такое гомофильное связывание играет важную роль в формировании тканей в процессе развития. Кадгериновые белки образуют димеры, которые связываются друг с другом.
Показаны три типа взаимодействия кадгеринов.
Прямые измерения силы взаимодействия показывают,
что наибольшей степени перекрывания соответствует максимальная прочность связывания.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
К основным факторам патогенности (вирулентности) относят способность микроорганизмов к колонизации, их устойчивость к разным микробицидным факторам организма, свойства инва-зивности и токсигенности, а также способность к длительному персистированию.
Способность к колонизации. Адгезия. Факторы колонизации.
Размножению бактерий в первичном очаге инфицирования предшествует адгезия [от лат. adhaesio, прикрепляться к чему-либо], то есть закрепление бактерий на поверхности клеток, что, собственно, и служит началом инфекционного процесса. Прикрепление к поверхности клеток (например, к эпителию слизистых оболочек) обеспечивают адгезины, или факторы колонизации — различные микробные продукты — молекулы адгезии (белки, ЛПС, липо-тейхоевые кислоты). Молекулы адгезии могут располагаться непосредственно на поверхности бактериальной клетки либо входить в состав микроворсинок или капсул.
Взаимодействие инфекционного агента с эпителиальными клетками происходит в результате нескольких типов связей, различных по природе и специфичности. Выделяют связи, основанные на взаимодействии электростатических сил, обусловленные гидрофобными свойствами поверхности, лиганд-рецепторные взаимодействия.
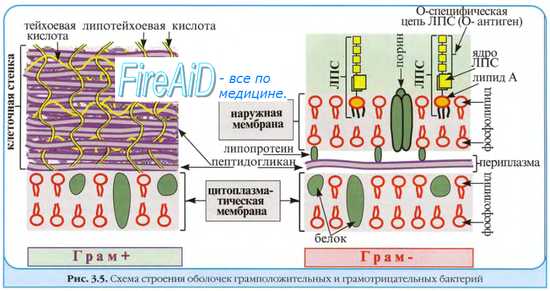
Заряд. Бактериальные и эукариотические клетки заряжены отрицательно, но поверхностные микроворсинки грамотрицательных бактерий снижают заряд бактерий и уменьшают электростатические силы отталкивания.
Гидрофобность. Бескапсульные бактерии обладают высокой гидрофобностью, усиливающей адгезивность; гидрофобные участки обладают сродством к лигандам на поверхности эукарио-тических клеток, что и приводит к прочности связи.
Специфические взаимодействия. На поверхности бактерий имеются молекулы, способные к стереоспецифичному связыванию с комплементарными молекулами на мембранах эукарио-тических клеток (например, гемагглютинины или тейхоевые кислоты).
Другие механизмы колонизации. Некоторые бактерии способны «заранее подготавливать» место для дальнейшего размножения; например, нейраминидаза облегчает проникновение холерного вибриона через слой слизи и контакт с сиалосодержащими рецепторами эпителия кишечника. Микроорганизмы также способны сорбироваться на бактериях, уже колонизировавших поверхность слизистых оболочек, либо связывать белки (например, фибронектин), рецепторы к которому имеются на многих клетках макроорганизма. У капсулированных бактерий в прикреплении активно участвуют полисахариды капсулы. Для успешной колонизации очага первичного инфицирования бактерии должны выдержать действие многочисленных и разнообразных микробицидных факторов хозяина. Для защиты от них микроорганизмы активно используют ряд структур (например, капсулы) и синтезируемых веществ (например, ферменты).
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

Грибы рода Candida являются уникальными микроорганизмами, имеющими огромный диапазон адаптационных возможностей (от сапрофитов до комменсалов и оппортунистов), позволяющих им успешно проживать как в различных анатомических местах организма хозяина, так и во внешней среде. И хотя этим грибам и вызываемым ими инфекциям посвящено большое количество работ, многие вопросы остаются до конца не решенными, например, такой «простой» вопрос, как происхождение источника инфекции при кандидозе. Считается, что большинство штаммов, выступающих в качестве причины заболевания, являются комменсалами самих пациентов. Возникает вопрос, каковы причины и вклад грибов рода Candida при переключении от «безобидного» сосуществования до агрессии? И одним из параметров, характеризующих трансформацию биологических свойств грибов рода Candida при смене условий существования (внешняя среда/человек), является способность к адгезии на эпителиоциты человека.
В тоже время, возрастающая частота кандидозов, в т.ч. вызванных Candida non albicans, может свидетельствовать об импорте инфекции и (или) о замене, в силу каких-то обстоятельств, «добропорядочных» комменсалов на агрессивные («дикие») штаммы.
Целью данного исследования было сравнить природные и клинические изоляты грибов рода Candida:
- по чувствительности к антифунгальным препаратам in vitro;
- по адгезивной способности в системе «Candidaspp. - буккальные/вагинальные эпителиоциты» in vitro.
1. Исследуемые культуры. В работе были использованы:
- культуры грибов рода Candida, выделенные из: а) влагалища женщин с различной генитальной патологией (n = 22); б) ротовой полости больных с кандидозом ротовой полости (n = 12); в) окружающей среды (n = 10);
- штамм CandidaalbicansATCC 10231.
Все клинические изоляты первоначально идентифицировали с помощью теста на ростовую трубку. Candida non albicans виды идентифицировали далее с помощью тест-системы «Auxocolor» (BioRad, Франция). Среди штаммов, выделенных из влагалища, одна культура была идентифицирована как Candida krusei, три - Candida glabrata; остальные - Candida albicans. Оральные изоляты все были представлены видом C. albicans. Природные штаммы были любезно предоставлены кафедрой биологии почв факультета почвоведения МГУ. Среди них было восемь коллекционных (три культуры - Candida guilliermondii, три - Candida tropicalis, одна - Candida maltosa, одна - Rhodotorula rubra) и две музейные культуры. Музейные культуры были реидентифицированы нами как Rhodotorula rubra и Candida kefyr («Auxocolor»).
2. Чувствительность к антифунгальным препаратам. Для определения чувствительности грибов использовали коммерческую тест-систему «Fungitest» (BioRad, Франция), включающую следующие препараты:
- Флуконазол в концентрации 8 и 64 мкг/мл,
- Итраконазол 0,5 и 4 мкг/мл,
- Кетоконазол 0,5 и 4 мкг/мл,
- Миконазол 0,5 и 8 мкг/мл,
- Амфотерицин В 2 и 8 мкг/мл,
- 5-флюороцитозин 2 и 32 мкг/мл.
Методику проводили согласно прилагаемой инструкции.
Кроме этого, из тех лунок, где результат был расценен как «чувствителен», для оценки эффекта действия на грибы антифунгального препарата, производили высев 10 мкл содержимого каждой лунки микропланшета на среду Сабуро. Если через 48 часов инкубации при 28°С проявлялся рост грибов, то эффект расценивали как фунгистатический; если роста не было - как фунгицидный.
Результаты. Природные изоляты грибов рода Candida проявили наибольшую чувствительность к антифунгальным препаратам (77,3% чувствительных штаммов) по сравнению с клиническими (59,8%). Максимальную активность в отношении природных и клинических изолятов грибов рода Candida продемонстрировали амфотерицин В (100% чувствительных штаммов) и 5-флюороцитозин (100% среди природных по отношению к 77-92% клинических). Природные штаммы, в отличие от клинических изолятов, показали высокую чувствительность кетоконазолу, но оказались устойчивыми по отношению к миконазолу в концентрации 0,5 мкг/мл (при концентрации 8 мкг/мл показали 100% чувствительность). Обращает внимание низкая чувствительность клинических изолятов к флуконазолу при обеих концентрациях. Для всех антимикотиков было отмечено, что если при минимальной концентрации препарат оказывал фунгистатическое действие, то при увеличении концентрации - эффект становился фунгицидным.
Штамм C.albicans АТСС 10231 продемонстрировал 100% чувствительность ко всем антимикотикам, входящим в набор «Fungitest».
Вывод. Природные изоляты проявили более высокую чувствительность к антифунгальным препаратам, чем клинические, что, возможно, является отражением общей тенденции роста числа резистентных к антифунгальным препаратам клинических штаммов грибов рода Candida.
3. Адгезия к эпителиальным клеткам здорового человека in vitro. Для изучения адгезивной способности грибов было выбрано две модели: буккальный и вагинальный эпителий. Известно, что успешность адгезии (с последующей колонизацией слизистых поверхностей человека) зависит от координированной регуляции большого количества генов и тестирования множества окружающих параметров, включая физиологические, экологической ниши заселения. Исходя из сказанного, адгезия проводилась с созданием следующих условий: температура 37°С, контактный эпителий из ротовой и вагинальной полости здоровых лиц, pH культуральной среды соответствовал физиологическим показателям той экологической ниши, из которой был получен эпителий (адгезию к вагинальному эпителию для всех штаммов проводили при рН 4,0-4,5; к буккальному - при рН 7,0).
Адгезивная способность всех изученных культур грибов рода Candida оценивалась на буккальном эпителии. У части культур (17 вагинальных, 7 оральных и 5 природных) адгезия оценивалась одновременно на буккальном и вагинальном эпителии.
С целью определения индекса адгезии (ИА) смешивали равные объемы эпителия и грибов при соотношении клеток 1:100 соответственно. Пробирки встряхивали в ротаторе 1 час при 37°С, 70 об/мин. Затем готовили мазок, который окрашивали метиленовой синью. Подсчитывали индекс адгезии - среднее количество адгезированных грибов в пересчете на один эпителиоцит. Определение индекса прочности адгезии (ИПА) проводили следующим образом - эпителиоциты с прикрепившимися кандидами наслаивали на фиколл-верографин (плотность 1,077 г/см 3 ) и центрифугировали при 1000 об/мин 10 минут. Из клеточного осадка делали мазок и подсчитывали индекс прочности адгезии (количество кандид в пересчете на один эпителиоцит, оставшихся адгезированными после прохождения через фиколл-верографин). Процент прочноадгезированных кандид по отношению ко всем прикрепившимся определяли по формуле: ИПА/ ИА × 100%.
Статистическую обработку полученных результатов проводили, используя критерий Стьюдента.
Результаты. Сравнение индексов адгезии оральных, вагинальных и природных изолятов грибов рода Candida к буккальному эпителию (таблица 1) показало наличие достоверно более низкого показателя у природных изолятов по отношению к клиническим штаммам (p<0,01). В то же время, отношение индекса прочности адгезии к индексу адгезии было достоверно выше среди природных изолятов по сравнению с оральными и вагинальными (p<0,05).
Штамм C.albicans АТСС 10231 и природные изоляты имели практически равный индекс адгезии (3,9 и 3,8 соответственно) к буккальному эпителию, но количество прочноадгезированных кандид по отношению ко всем адгезированным клеткам у АТСС штамма было в два раза ниже (25,6% и 50,8% соответственно).
Сравнение индексов адгезии к вагинальному эпителию показало наличие более высокого показателя у вагинальных изолятов в сравнении с оральными (p<0,02) и природными. В то же время, прочность крепления к вагинальному эпителию изолятов из разных экологических ниш достоверно не различалась.
- Способность вагинальных изолятов Candida albicans одинаково успешно осуществлять адгезию к клеткам вагинального и орального эпителия (одинаковые индексы адгезии и индексы прочности адгезии) позволяет предположить, что вагинально-оральный путь передачи штаммов рода Candida в родах новорожденному является эволюционно-отраженным физиологическим путем создания нормобиоты ротовой полости, но может стать путем патофизиологическим (т.е. путем развития орального кандидоза у новорожденных) в условиях клинически выраженного вульвовагинального кандидоза у матери.
- Хорошо выраженная сайт-специфическая адгезия оральных изолятов Candida albicans к буккальному эпителию может свидетельствовать об успешном импорте оральной кандидозной инфекции в госпитальных условиях (через предметы личного пользования: ложка, стакан и т.д.), особенно когда речь идет о заселении ротовой полости новорожденного или грудного младенца.
- Можно предположить, что существенный процент заболеваний (и оральных, и вагинальных кандидозов) может быть связан с частыми орально-генитальными контактами, а источником кандид могут служить как ротовая, так и вагинальная (более вероятно) полости сексуальных партнеров.
- В связи с тем, что природные штаммы демонстрируют высокую прочность крепления к буккальному эпителию, можно высказать предположение, что при наличии определенных условий природные изоляты имеют шанс стать успешными оральными комменсалами.
Таблица 1. Сравнение адгезивной способности природных и клинических изолятов грибов рода Candida к буккальному и вагинальному эпителию
Читайте также: